Review Article
Open Access, Volume 3
A Narrative Review on Synaptotagmin 13
Stefanie J Willmann*
Technical University of Munich, D, 85748 Garching b. Munich, Germany.
Technical University of Munich, D, 85748 Garching b. Munich, Germany.
Synaptotagmin 13 (hereinafter stated as Syt13) is a pro-endocrine factor required for endocrine-lineage specification during mouse pancreatic development. To notion, activation of the transcription factor Neurog 3 (hereinafter stated as Ngn3) epithelial cells in the pancreatic cord leads to the rapid progression into post-mitotic Ngn3high endocrine -committed precursor state. Through co-expression within an micro-environmental niche of Syt13 along with Ngn3, we challenged the hypothesis by postulating a model in which a particular level of expression will re-semble to give rise to cells, inherently committed to an endocrine precursor state.
In embrogenesis, a diverse set of signals and factors determine the onset of the particular organ formation. A detailed roadmap of these processes and machinery involved in lineage segregation of a formerly multi-potent state of cells is still missing. The gene product of Syt13 suggests to be involved in early embryonic development, hence expression within adult tissues is not discussed. Syt13, a Ca2+-independent atypical Synaptotagmin-family member, concomitant with Ngn3 indicates to be lineage decisive in the segregation of the neuro-endocrine progenitor state cells in a particular micro-environmental niche within the epithelial cord of the evolving pancreatic gland. Mechanistically, Syt13, as a vesicle trafficking protein, initiated exocystic transport within the filamentous actin network of a cell and re-sembles the exosomes on the inner membrane surface. Pre-liminary interactom analysis pinpoints to the association of the gene product of Syt13, concomitant with Ngn3 expression, of a highly regulated cellular process for exocystic release of signal and factors for cell-to-cell communication.
Keywords: Synaptotagmin-13; Ngn3; Exocystic release; Model proposition.
Manuscript Information: Received: Jul 31, 2023; Accepted: Aug 18, 2023; Published: Aug 25, 2023
Correspondance: Stefanie J Willmann, Technical University of Munich, D, 85748 Garching b. Munich, Germany.
Email: SJWillmann@gmx.de
Citation: Stefanie JW. A Narrative Review on Synpatotagmin 13. J Oncology. 2023; 3(2): 1104.
Copyright: © Stefanie JW 2023. Content published in the journal follows creative common attribution license.
Syt13 expression levels initiates different pathways within a cells
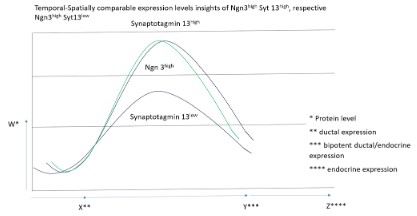
In a global gene expression profiling during mouse pancreatic development, Syt13 expression pattern was previously identified and characterized into Syt13low and Syt13high. To further evaluate the different expression patterns of the gene product of Syt13, correlation in expression to the Tf Ngn3 could emphasize the role of Syt13 in lineage dcisive segregation processes during mouse embryogenesis. In a postulazed model, the temporal-spatially expression level of Syt13high Ngn3high versus Syt13low Ngn3high is taken into consideration under the highlight of the pathway analysis (Figure 1). Consistent with current findings, precursor states in lineage segregating cells express Syt13, along with a distinct subset of other gene products, here concomittant with the neuroendocrine initiating factor Ngn3. Neuro-endocrine factor Ngn3 is well described in the context of connecting the cell cycle and pancreatic endocrine lineage-specification. Notably, Ngn3 expression level are separated between Ngn3high versus Ngn3low, indicating for the Ngn3high micro-environmental niche a partial decision of endocrine lineage decision, whereas the Ngn3low cell micro-environmental niche implicates to pre-empts the endocrine-commitment.
Below, we discuss potential conncetions between the cell cycle and (related) Syt13 [1,2], respective Ngn3, gene product levels in endocrine-committed cells, emerging out of the micro-environmental niche, here stated as endocrine birth niche. Understanding, how cycling Syt13/Ngn3 progenitors integrate their behaviour within surrounding dynamics of developing epithelial micro-environmental niche will be important. For example, what determines a progenitor maintaining state and/or symmetric endocrine birth divisions, and the end-fate endocrine potential of Syt13 expressing progenitors, could lead to better disease understanding, through bypassing the essential network, inherently necessary for proper organ formation [3-6].
Gain and loss of functional studies show the impact of Ngn3 in instructing endocrine lineage segregation, respective differentiation, but other studies had already begun to suggest correlations between a more finely tuned spatial-temporal gene product expression level with Syt13 and the potential of the Syt13high Ngn3 sub-population to produce endocrine cells (Figure 1). Bahkti et al. (2022) reported that a high expression levels of Syt13 in endocrine precursor cells prime the ß-cell fate, contrary, lack of Syt13 reduces ß-cell specification, futher indicationg that the mitotic index in the endocrine-biased cells determine the the expense of endocrine committed cells. In addition to high-level Ngn3 expression within the endocrine-biased mitotic cell population, Syt13 downstream target is expressed at different protein product levels, suggesting that endocrine-biased cells undertake a concerted effort to build the framework of a gene regulatory network that primes intra-epithelial Syt13low progenitors to the endocrine lineage, before their commitment. Such concepts have been referred to as lineage-priming and is familiar to concepts in the thematic field of neurological and intestinal stem cells. We propose, that the Syt13lowNgn3 state represents a mitotic metastable progenitor state in which low Syt13 protein product levels, under normal conditions, begin to drive cells towards the endocrine lineage (Figure 1 and Figure 2) [7-10].
The notion, that Ngn3 can maintain a metastable progenitor state at low gene product expression level, but at higher gene product expression levels trigger cell-cycle exit and endocrine lineage commitment is a concept not without precedent. In neural progenitors, the Neurog family member Neurog 2 is actively phosphorylated by DdKs during S-G2-M phases, resulting in a low level unstable form that activates progenitor associated target genes. During G1, however, the increased acitivity of Cdk-inhibitors results in accumulation of underphosphorylated Neurog2, which is stabilized and targets different suites of genes that promote neural differentiation [11,12].
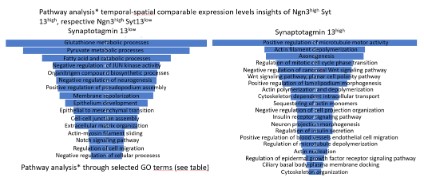
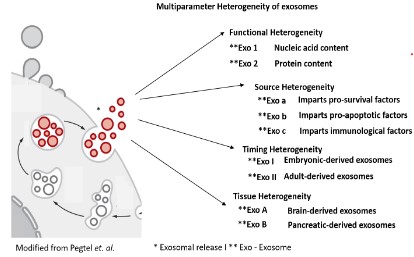
The pathway analysis underpins the association of Syt13, along with Ngn3, in cytokinetic processes of a single cel (Figure 2). Mainly, the Syt13low Ngn3high sub-population refers to Gene Ontology (hereinafter stated as GO) terms such as negative regulation of neurogenesis, contrary, the sub-population the Syt13high Ngn3high illustrates as GO terms a sub-set of genes involved in Axonogenesis and neuron projection morphogenesis. Initiative, the Syt13low Ngn3high sub-population, positively regulates pseudopodium association, stepwise delamination might occur in the sub-population of Syt13high Ngn3high as pathway analysis reflects a high abundance with the gene regulatory network in processes associated to the GO terms positive regulation of microtubule motor activity, actin filament depolymerization, regulation of mitotic cell cycle phase transition, positive regulation of lamelipodium morphogenesis and cytoskeleton dependent intra-cellular transport (Figure 2). In a further step, interaction partners, referred to the gene products ofSyt13, highlight the association of membrane trafficking proteins and hint to the role of the gene products of Syt13 in the process of exocyst function. We further postulate a model for precise analysis of expression pattern and localization of the gene products of Syt13 in defining the cellular processes of involvement of the gene product of Syt13 (Figure 3) [13-17]. We suggest, that the gene product of Syt13 acts as a inner peripheral membrane protein in exosome shuffling, thus aiming in exocytotic processes as lipid anchor of the inner membrane proteins of the inner cell surface of a single cell. The multiparameter heterogeneity of exosomes is well described in other contextes, but deciphering functional heterogeneity, source heterogeneity, timing heterogeneity and functioal heterogeneity will aid the understanding of multicellular life.
Here we present a roadmap to further enhance localization, function and regulatory netonetwork regarding the gene products of Syt13. Notably, the onset of Ngn3 expression triggers lineage segregation, thus concomitant to the Syt13 expression profile, lineage priming to endocrine precursor state is a temporal and spatial window. In the aspect of cell-to-cell communication, the Golgi apparatus serves as regulator of protein trafficking, releasing vesicle into the cytoplasm from the endosomal membrane. As Stipulated, Syt 13 acts as a trans-acting modulator of exosome release, that mediates intracellular protein trafficking pathways [18,19].
The close proximity to Rab exosome effector proteins, underpins the role of Syt13 in the exocytotic machinery. Further studies, related to the exosome content, as well as the exosome itself as remodeler of the extracellular matrix after fusion of the vessel will faciliate disease management. In a perspective, the involvement of Syt13 in exosome biogenesis invites to numerous investigative quests, likely such as how cell-to-cell communication of different cells apportion exosome biogenesis between their endosome and plasma membranes, regulate biogenesis and identify the mechanism that mediate functional delivery of bio-active proteins. Lipids, and nucleic acids into target cells [20,21].
On might question the scientific content therein, nonetheless exosome-mediated signaling plays a role in PD-L1 signaling in immunosuppression and in transfer of, f.e. EGFRvIII in cancerogenesis. Thus, although one should be skeptical when considering whether exosomes are important for a particular biological process, this skepticism should be tempered by the high likelihood that multicellular life evolved in the context of constant, neverending formation and uptake of exosomes. In conclusion, further studies on expression and localization of the gene product Syt13 will enhance current findings in the field physiological processes (e.g. development, tissue homeostasis, aging, metabolic regulation, stress, circadian rhythms), numerous non-infectious diseases (e.g. cancerogenesis, inflammation, metabolic disorders, neuro-degeneration, auto-immunity)and a similar broad array of infectious diseases (e.g. those caused by infection with viruses, protozoans, fungi, ect.).
Publicly available datasets:
https://github.com/theislab/2022_Bakhti_pancreas_Syt13Statistical analysis: All statistical analysis was performed with Excel and presented in Figure legends.
Highlights
- cytoskeletal rearrangement is initiated through Synaptotagmin-13 (Syt 13).
- Syt13high versus Syt13low inititiates a diverse segregation within the epithelial cord in mouse embryogenesis.
- Neuro-endocrine Transcription Factor (TF) Neurog 3 (Ngn3) concomittant with Syt13 regulates delamination within a micro-environmental niche.
- misfunction of Syt13 leads to severe metabolic diseases, cancerogenesis and cerebral diseases.