Introduction
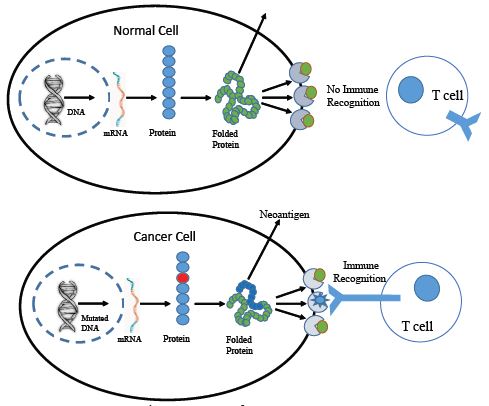
Cancer is a major concern of mortality and morbidity worldwide, with a large economic burden on the world’s population. Tumour heterogeneity restricts the concept of a single drug that can be universally effective against all cancer types. Different treatment approaches, such as chemotherapy, radiotherapy, and immunotherapy, have been implemented in the treatment of cancer, but very few have been identified without any side effects. Among these breakthroughs, the discovery and exploration of cancer neoantigens stand out as a potential game-changer in the field of cancer immunotherapy. Neoantigens are mutated proteins that arise from somatic mutations in cancer cells. These mutations can result from errors during DNA replication, exposure to carcinogens, or due to other genetic abnormalities. Neoantigens are unique to each patient or they may be common in a specific type of cancer but are not present in normal cells, making them attractive targets for the immune system to recognize and attack. Neoantigens are derived from various types of genetic alterations, including single nucleotide variations, insertions, deletions, alternative splicing and gene fusions. They can be classified into two main categories: personalized neoantigens, which are unique to an individual patient’s tumor, and shared neoantigens, which are present in multiple patients with the same type of cancer. Cancer neoantigens have emerged as a new horizon in cancer research such as Programmed Death-1 (PD-1) and Cytotoxic T Lymphocyte-Associated Antigen-4 (CTLA-4). Various Tumor Specific Antigens (TSA) have been identified as markers only. A few TSAs have been established as cancer treatment targets, like Human Epidermal growth factor Receptor 2 (HER2), Eoestrogen Receptors (ER), Progesterone Receptors (PR), etc. TSAs therefore provide a promising approach to directing cancer treatment without any collateral damage to normal body cells. Furthermore, only a few of them have been used in early-stage cancer diagnostics.
Labyrinthin is unique among TSAs because it is exclusively expressed in adenocarcinoma-type cancer cells and not in other cancers, and is not found to exist in normal cells. Our recent article showed that labyrinthin is present exclusively on the surface of non-permeablized adenocarcinoma cells. Whereas, in permeablized cells likely exhibit co-recognition of intracellular ASPH proteins along with internalized labyrinthin. Thus, labyrinthin not only provides better treatment specificity to target only adenocarcinomas, but it could also be a promising early cancer marker that can be used in companion diagnostics. Herein, we review the latest and most promising cancer marker targets such as Labyrinthin, as well as other targets in regards to their specificity and scope in cancer therapeutics.
The significance of cancer neoantigens
Cancer neoantigens have garnered considerable attention due to their potential therapeutic implications that include:
(1) Precision immunotherapy: Traditional cancer treatments like chemotherapy and radiation therapy often result in collateral damage to healthy cells. Neoantigens offer a more precise way to target cancer cells while sparing normal tissues, thus minimizing side effects.
(2) Personalized medicine: The uniqueness of neoantigens in each patient’s tumor means that treatment strategies can be tailored to the individual. This approach has the potential to improve treatment efficacy by enhancing the immune system’s ability to recognize and eliminate cancer cells.
(3) Overcoming immune evasion: Cancer cells often employ various mechanisms to evade the immune system’s surveillance. Neoantigens can help bypass these evasive strategies by presenting the immune system with novel targets that have not been previously encountered.
Harnessing neoantigens for immunotherapy
ential of cancer neoantigens in immunotherapy to include: Personalized neoantigen vaccines are designed based on the genetic makeup of a patient’s tumor. These vaccines stimulate the immune system to recognize and attack cancer cells displaying specific neoantigens. Early clinical trials have shown promising results, with evidence of immune activation and tumor regression in some patients. T cells are extracted from a patient’s blood, genetically modified to target specific neoantigens, and then infused back into the patient. This approach has demonstrated success in certain cases of advanced melanoma and other cancers.
Checkpoint inhibitors: Drugs that block proteins that dampen the immune response. Combining these inhibitors with neoantigen-targeted therapies are expected to enhance the immune system’s ability to recognize and eliminate cancer cells. Essentially, creating an adjunctive cancer treatment with both an offensive and defensive approach.
Emerging cancer neoantigens
KRAS G12D
A mutation in the KRAS gene is frequently found in various cancers, such as pancreatic, colorectal, and lung cancers. The G12D mutation leads to the expression of a neoantigen that has been studied for its potential immunotherapeutic applications. The KRAS gene, a critical player in cellular signalling pathways, is among the most frequently mutated genes in human cancers. Within the realm of cancer genetics, one specific KRAS mutation has gained particular attention: KRAS G12D. This mutation has profound implications for cancer development, progression, and potential therapeutic interventions. The KRAS gene encodes a small GTPase protein that plays a central role in transmitting signals from cell surface receptors, such as growth factor receptors, to intracellular signalling pathways that regulate cell growth, differentiation, and survival. Activation of KRAS triggers downstream cascades that control key cellular processes. However, when KRAS becomes mutated, these pathways can be dysregulated, contributing to the initiation and progression of cancer. KRAS G12D mutation act as a driver of oncogenesis which involving a single nucleotide change from G to A at position 12 in the KRAS gene. This mutation leads to the substitution of glycine (G) with aspartic acid (D) at amino acid position 12 in the KRAS protein. This seemingly small alteration has profound consequences for cellular function and cancer development. The G12D mutation results in a constitutively active KRAS protein that is locked in the GTP-bound state, preventing its normal regulation and rendering it continuously active. This hyperactive KRAS protein drives uncontrolled cell growth, survival, and proliferation, contributing to the development and maintenance of various cancer types. The KRAS G12D mutation has been implicated in a range of cancers, including but not limited to the following cancers such as in colorectal cancer KRAS G12D mutations are frequently found can influence disease progression and response to treatment. As like, in pancreatic ductal adenocarcinoma often harbours KRAS G12D mutations, contributing to the aggressive nature of this cancer. In non-small cell lung cancer, KRAS G12D mutations are associated with a distinct clinical profile and therapeutic challenges. Beside these KRAS G12D mutations have been detected in ovarian, gastric, and other cancer types, showcasing their broad significance in oncogenesis.
Therapeutic challenges with KRAS G12D
The presence of the KRAS G12D mutation has posed challenges for cancer treatment, as it was traditionally considered «undruggable» due to the complex nature of targeting GTPases directly. However, recent advancements in drug development and molecular understanding have opened up new possibilities still the KRAS G12D mutation exemplifies the intricate interplay between genetics, signalling pathways, and cancer development. Its prevalence across diverse cancer types underscores its significance in oncogenesis. While the KRAS gene and its mutations, including the well-known KRAS G12D mutation, have attracted significant attention in cancer research, there are several limitations and challenges associated with targeting KRAS mutations as neoantigen targets for cancer therapies. Here are some of the key limitations. The complex signalling network involving KRAS mutations poses challenges in developing effective targeted therapies. Dysregulation of multiple downstream pathways can occur, making it difficult to predict the precise consequences of inhibiting a specific KRAS mutation [1].
Resistance to targeted therapies is a significant concern in the field of oncology. Cancer cells can develop alternate pathways to bypass KRAS signalling inhibition, rendering therapies less effective over time [2]. Further study suggests that it lacks “Druggable Binding Pockets” that the structural characteristics of mutant KRAS proteins pose challenges in developing small molecule inhibitors that can effectively bind to and block their activity [3]. Another concern with KRAS G12D is that the immunogenicity of KRAS mutant neoantigens varies across different patients and cancer types, limiting the efficacy of immune-based therapies targeting these many distinct mutations [4]. Furthermore, tumor Heterogeneity in KRAS mutant allele frequency and expression can impact the effectiveness of targeted therapies and contribute to treatment resistance [5]. Despite extensive research, there has been limited clinical success in developing therapies that effectively target KRAS mutations, which has highlighted the challenges in translating preclinical findings to the clinic [6].
BRAF V600E
The BRAF V600E mutation is common in melanoma and certain other cancers. The discovery of driver mutations in cancer has revolutionized our understanding of tumorigenesis and paved the way for targeted therapies. Among these, the BRAF V600E mutation has emerged as a prominent player, offering insights into oncogenesis and treatment strategies in various cancers. The neoantigen resulting from the BRAF V600E mutation has been explored as a target for immunotherapy. However, while its significance is undeniable, limitations also exist, and shaping the clinical landscape. The BRAF gene encodes a serine/threonine kinase involved in the MAPK/ERK signalling pathway, regulating cell growth and proliferation. The V600E mutation, resulting in a constitutively active BRAF protein, has been extensively studied across multiple cancer types. In melanoma, the BRAF V600E mutation is a hallmark alteration, present in about 50% of cases. It drives hyper activation of the MAPK pathway, contributing to uncontrolled growth. Targeted therapies like vemurafenib and dabrafenib, specific to BRAF V600E, have shown remarkable clinical responses in melanoma patients, illustrating the mutation’s role as both a diagnostic marker and therapeutic target [7]. The BRAF V600E mutation is also associated with a distinct molecular subtype of colorectal cancer characterized by poor prognosis, resistance to standard therapies, and enhanced tumor invasiveness [8]. It coexists with other mutations, emphasizing the complex genetic landscape of colorectal cancer. Beside these, in papillary thyroid carcinoma, the BRAF V600E mutation has prognostic significance, correlating with aggressive features and an increased risk of recurrence [9]. It has led to investigations into targeted therapies for this subtype of thyroid cancer. Moreover, BRAF V600E is prevalent in hairy cell leukaemia, a rare B-cell malignancy [10].
Targeted therapy with BRAF inhibitors has transformed the treatment landscape for this disease. The major limitations related to BRAF V600F are 1) Tumor Heterogeneity- Cancer is inherently heterogeneous, with tumor cells harbouring diverse mutations. The role of BRAF V600E can be modulated by coexisting alterations, influencing disease progression and treatment responses. 2) Resistance to Targeted Therapies- Despite initial successes of BRAF inhibitors, resistance mechanisms inevitably emerge. MAPK reactivation through alternative pathways, such as MEK, can render targeted therapies ineffective [11]. 3) Cancer Type Specificity- While the significance of BRAF V600E is evident in certain cancers, its prevalence and relevance vary across tumor types, necessitating a tailored approach to diagnosis and treatment. Beside all these, the mutation’s frequency varies among different populations, affecting its clinical relevance and impact on therapeutic decisions. The BRAF V600E mutation’s role in oncogenesis is substantial, offering insights into targeted therapies and personalized medicine. However, its limitations, including tumor heterogeneity and therapeutic resistance, underscore the complex nature of cancer biology. Advances in understanding the mutation’s context-dependent effects will refine treatment strategies, emphasizing the importance of comprehensive molecular profiling and multidisciplinary approaches in managing BRAF V600E-driven cancers.
EGFRvIII
The epidermal growth factor receptor variant III (EGFRvIII) is a truncated and constitutively active mutant of the epidermal growth factor receptor (EGFR) that lacks exons 2-7 due to an in-frame deletion. This mutation is commonly observed in various cancers, especially Glioblastoma Multiforme (GBM), a highly aggressive brain tumor. EGFRvIII is frequently found in GBM, where it contributes to tumorigenesis and treatment resistance. It promotes cell proliferation, invasion, and survival through dysregulated signalling pathways, making it an attractive therapeutic target. Strategies to target EGFRvIII include monoclonal antibodies, small molecule inhibitors, and immunotherapies [12]. Beside these EGFRvIII can also be used in targeted Immunotherapies; for example, the tumor-specific nature of EGFRvIII’s offers potential for immunotherapies. Vaccines targeting EGFRvIII have been developed, aiming to trigger an immune response against tumor cells expressing this mutant receptor [13]. Chimeric Antigen Receptor (CAR) T-cell therapy targeting EGFRvIII has also shown promise in preclinical studies [14]. Beside the clinical significance there are some major limitations with EGFRvIII. For example EGFRvIII expression is not uniform across cancer cells within a tumor, making it challenging to target all cells effectively. This heterogeneity contributes to treatment resistance and disease recurrence [15]. Moreover, Tumor can develop resistance mechanisms to therapies targeting EGFRvIII. Up regulation of alternative signalling pathways or mutations in downstream effectors can circumvent the effects of EGFRvIII inhibition [16]. Other than these, it has been found that the tumor microenvironment, especially in GBM, can be highly immunosuppressive, limiting the effectiveness of immunotherapies targeting EGFRvIII [17]. One study reported that immune responses against EGFRvIII may be thwarted by antigenic variability due to tumor heterogeneity and the generation of antigenic escape variants [18]. Finally, with respect to brain tumors like GBM, effective drug delivery to the tumor site remains a challenge. The blood-brain barrier and tumor microenvironment restrict the penetration of therapeutic agents targeting EGFRvIII [19].
In conclusion, EGFRvIII’s prominent role in cancer progression has positioned it as a promising target for therapies, particularly in glioblastoma multiforme. While its tumor-specific nature presents unique advantages, the limitations, including heterogeneity, resistance mechanisms, and immunosuppression, necessitate comprehensive approaches that combine targeted therapies with strategies to overcome the identified obstacles. Continued research and clinical trials are essential to develop effective therapies that leverage EGFRvIII’s potential while addressing its limitations.
MAGE-A3
MAGE-A3 (Melanoma Antigen-A3) is a member of the antigen family of cancer-testis antigens, which are normally expressed but can also be aberrantly expressed in various cancers. MAGE-A3 has gained attention as a potential target for cancer immunotherapy due to its restricted expression pattern in cancer cells and role in tumor progression. MAGE-A3 is normally not expressed in healthy adult tissues, except for testis and placenta. However, it is frequently expressed in various cancer types, such as lung, melanoma, oesophageal, and bladder cancers. The overexpression of MAGE-A3 in cancer cells has been implicated in promoting tumor cell proliferation, resistance to apoptosis (programmed cell death), and evasion of immune surveillance [20-22]. The selective MAGE-A3 expression makes it an attractive target for cancer immunotherapy. Several therapeutic approaches have been explored to target MAGE-A3 for cancer treatment: (1) vaccines- MAGE-A3 has been investigated as a target for cancer vaccines. Cancer vaccines aim to stimulate the immune system to recognize and attack cancer cells expressing specific antigens like MAGE-A3 [23]. Immunotherapeutic approaches, such as immune checkpoint inhibitors, have been explored in combination with targeting MAGE-A3 to enhance the anti-tumor immune response [24]. Monoclonal antibodies targeting MAGE-A3 may be used to deliver cytotoxic agents directly to cancer cells expressing MAGE-A3, leading to targeted cell death [25]. Gene therapy approaches, such as using viral vectors to deliver therapeutic genes to cancer cells, have been explored for targeting MAGE-A3 [26].
While targeting MAGE-A3 holds promise for cancer immunotherapy, there are several limitations and challenges such as MAGE-A3 expression not being uniform across all patients with the same cancer type. This variability can limit the effectiveness of MAGE-A3-targeted therapies since only patients with MAGE-A3-positive tumors would benefit [27]. Tumors can also develop mechanisms to evade immune responses, such as down regulating MAGE-A3 expression or suppressing immune cells, which could reduce the effectiveness of MAGE-A3-targeted therapies [28]. Since MAGE-A3 is also expressed in the testis, there is a concern that immune responses against MAGE-A3 could lead to autoimmune reactions targeting the testis [29]. Finally, clinical trials targeting MAGE-A3 have shown variable results, with some trials failing to demonstrate significant improvements in patient outcomes.
CTAG1B
CTAG1B, also known as NY-ESO-1 (New York Esophageal Squamous Cell Carcinoma-1), is a cancer-testis antigen that was originally identified in esophageal cancer. It is a member of the MAGE (Melanoma Antigen) family of antigens. CTAG1B is normally not expressed in most adult tissues but is present in the testis and placenta. However, it is frequently expressed in various cancer types, including melanoma, lung, ovarian, and bladder cancers. CTAG1B plays a role in tumor progression and immune response evasion. Its expression in cancer cells, coupled with its limited expression in normal tissues, makes it an attractive target for cancer immunotherapy. It has been shown that CTAB is capable of interfering the efficiency of cancer therapy by regulation of cell metabolic reprogramming [30].
While CTAG1B-based therapies hold promise, they also have limitations such as Heterogeneity, Immune evasion, autoimmune responses, clinical efficacy etc. In heterogeneity, CTAG1B expression can vary across different cancer types and even within individual tumors. Not all patients with a particular cancer type express CTAG1B, limiting the potential patient population that could benefit from CTAG1B-targeted therapies [31]. Furthermore, tumors can develop mechanisms to evade immune responses, such as inhibiting the activation of T cells or expressing inhibitory molecules that counteract immune attacks targeted at CTAG1B [32]. Since CTAG1B is also expressed in the testis, there is a concern that immune responses against CTAG1B could lead to autoimmune reactions targeting normal testicular tissue [33]. Clinical efficacy is another concern, while CTAG1B-targeted therapies have shown promise in some clinical trials, their overall clinical efficacy may vary depending on the specific cancer type and patient population [34].
NY-ESO-1
NY-ESO-1 (New York Oesophageal Squamous Cell Carcinoma-1) is another cancer-testis antigen that has gained attention as a potential target for cancer immunotherapy. It was first identified in oesophageal cancer but has been found to be expressed in other cancer types, including melanoma, lung, ovarian, and bladder cancers. NY-ESO-1 is normally not expressed in most adult tissues but is present in the testis and placenta. NY-ESO-1 plays a role in tumor progression by promoting tumor cell growth, invasion, and metastasis. Its restricted expression in cancer cells and its immunogenicity make it an attractive target for cancer immunotherapy.
Several therapeutic strategies have been explored to target NY-ESO-1-positive cancers. Cancer Vaccines containing NY-ESO-1 peptides or DNA sequences encoding NY-ESO-1 have been developed to stimulate an immune response against cancer cells expressing NY-ESO-1 [34]. These vaccines aim to activate T cells to recognize and attack NY-ESO-1-positive cancer cells. Similar to the approach for MAGE-A3, adoptive T cell therapy involves engineering patient T cells to express receptors that specifically target NY-ESO-1 and then infusing these modified T cells back into the patients. These engineered T cells can recognize and eliminate NY-ESO-1-positive cancer cells [31]. Other than these, combination therapy approaches have been used are: Immune checkpoint inhibitors, such as PD-1 and CTLA-4 inhibitors, have been used in combination with NY-ESO-1-targeted therapies to enhance the effectiveness of the immune response against NY-ESO-1-positive tumors. Checkpoint inhibitors help remove cancer cell induced “brakes” on the immune system, thereby allowing T cells to more effectively identify and attack cancer cells.
While NY-ESO-1-targeted therapies show promise, they also have limitations. Some tumors may down regulate or lose NY-ESO-1 expression over time, which can lead to treatment resistance and reduced efficacy of NY-ESO-1-targeted therapies [35]. Tumors can develop mechanisms to evade immune responses, such as inhibiting the activation of T cells or expressing inhibitory molecules that counteract immune attacks. Not all cancer patients express NY-ESO-1, which limits the potential patient population that could benefit from NY-ESO-1-targeted therapies [36].
HER2
HER2 (Human Epidermal Growth Factor Receptor 2) is a protein receptor that belongs to the epidermal growth factor receptor (EGFR) family. It plays a pivotal role in cell growth, proliferation, and survival. Overexpression or amplification of the HER2 gene is associated with several cancers, most notably breast cancer. HER2-positive breast cancer is characterized by increased levels of the HER2 protein on the surface of cancer cells. HER2 has a significant role in cancer therapy. HER2-positive breast cancer can be treated with targeted therapies that specifically inhibit the HER2 protein’s activity. Notable HER2-targeted therapies include: Trastuzumab (Herceptin): A monoclonal antibody that binds to the HER2 protein, preventing its signalling pathways and leading to reduced cancer cell growth. In a slightly different approach Pertuzumab (Perjeta) is a monoclonal antibody that targets a different part of the HER2 protein, inhibiting HER2 heterodimerization with other HER family members and thereby likewise preventing HER2 receptor function. Moreover, Lapatinib (Tykerb) is a tyrosine kinase inhibitor that blocks the signalling pathways of HER2 and EGFR.
HER2-targeted therapies have revolutionized the treatment of HER2-positive breast cancer. They have significantly improved response rates, disease-free survival, and overall survival for patients with HER2-positive tumors. Patients with HER2-positive breast cancer receive HER2-targeted therapies, in addition to standard treatments, leading to more tailored and effective treatment approaches [37]. Despite their success, HER2-targeted therapies have limitations which are mentioned here. Some patients eventually develop resistance to HER2-targeted therapies, leading to disease progression [38]. Mechanisms of resistance are multifactorial and can involve alternative signalling pathways. Among the side effect of Trastuzumab and other HER2-targeted therapies can cause cardiac toxicity, particularly when used in combination with certain chemotherapies [39]. Regular cardiac monitoring is crucial during treatment. Moreover, HER2-targeted therapies are effective only for HER2-positive cancers (e.g., breast, lung, uterine etc.). Patients with HER2-negative tumors do not benefit from these treatments [40]. Beside these limitation, HER2-targeted therapies can be expensive, and their accessibility can be limited in certain regions or healthcare systems.
TP53
Mutations in the TP53 gene are common in a wide range of cancers. TP53 is a tumor suppressor gene that plays a critical role in regulating cell cycle progression, DNA repair, and apoptosis (programmed cell death). Mutations in the TP53 gene are among the most common genetic alterations in cancer. TP53 mutations can lead to loss of its tumor-suppressive functions, allowing cells to evade growth control and accumulate genetic damage, ultimately promoting cancer development. While TP53 itself is not directly targeted by therapies, its status has significant implications for cancer treatment. TP53 mutations are associated with poorer prognosis in various cancer types, as they can confer aggressive tumor behaviour and resistance to therapy. It has been found that TP53-mutated tumors often exhibit resistance to conventional cancer therapies, including chemotherapy and radiation therapy. Although not targeting TP53 directly, some therapeutic strategies aim to exploit vulnerabilities in TP53-mutated cancers by targeting related signalling pathways. Like other neoantigens, TP53-based cancer treatment also faces several challenges.
The most common challenge is related to mutations that are highly heterogeneous, with various mutation types leading to diverse functional consequences [41]. This complexity makes it challenging to develop targeted therapies against a wide range of TP53 alterations. Not all cancers harbour higher frequency of TP53 mutations; some cancers show have less frequencies TP53. Therefore, TP53-targeted therapies would be relevant only to TP53-mutated cases [42,43]. Another limitation related to TP53 is lack of direct targeting. Unlike other molecular targets, TP53 is a transcription factor involved in multiple pathways, making it difficult to develop traditional small-molecule inhibitors against it [44,45]. Furthermore, TP53’s role in multiple cellular processes adds to the complexity of developing therapies that specifically restore its functions without causing harmful side effects [44]. Explored in cancers like sarcomas and certain leukaemia types [46,47].
Labyrinthin
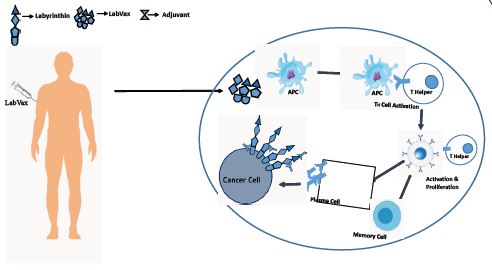
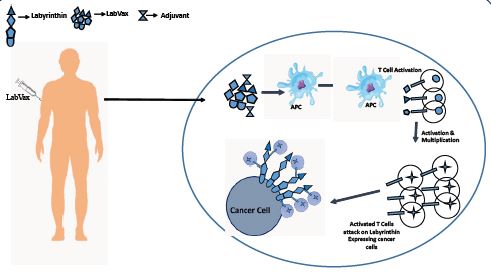
labyrinthin has emerged as a promising tumor specific antigen that has shown potential in the diagnosis and immunotherapy of various adenocarcinomas [48-57]. Research has shown that labyrinthin is consistently present in lung adenocarcinomas, leading to further investigations into its presence in other tumor types [57]. Multiple studies have demonstrated that labyrinthin is present in adenocarcinoma of the breast, bronchial tract, stomach, small intestine, pancreas, colon, and other glandular-related sites. This suggests that labyrinthin may have a role in sorting adenocarcinomas from other tumor types arising in various tissues. Given the specificity and selectivity of labyrinthin associated with adenocarcinoma and no other cancers or normal cells (eg., as opposed to HER-2 Neu) Numerous experimental approaches have supported the role of labyrinthin as a tumor specific antigen. One study utilized immunofluorescent labelling to demonstrate that the anti-labyrinthin mouse monoclonal antibody, MCA 44-3A6, binds to and persists on the cell surfaces of A549 adenocarcinoma cells [58].
Collectively, available data indicate that labyrinthin is accessible to the immune system and can be targeted for immunotherapeutic interventions. The development of labyrinthin-based treatments holds promise due to its ability to fulfil several criteria necessary for effective immunotherapeutic [59]. Firstly, the selectivity and convenient location of labyrinthin makes it an ideal target for immunotherapy, as it minimizes potential off-target effects on healthy tissues. In addition, labyrinthin is consistently present in adenocarcinomas derived from various tissues, making it a potential candidate for pan-adenocarcinoma immunotherapies.
The identification and characterization of neoantigens, such as labyrinthin, are rare but crucial for diagnostic and therapeutic purposes. Labyrinthin’s selective expression on the cell surface of adenocarcinomas, combined with its absence in normal and other cancer cell types, makes it a valuable marker for distinguishing adenocarcinomas from other tumor types. Furthermore, the presence of labyrinthin in a variety of adenocarcinomas derived from different tissues highlights its potential as a pan-adenocarcinoma target. The significance of labyrinthin as a tumor-specific antigen lies in its ability to accurately distinguish adenocarcinomas from other tumor types, regardless of the tissue of origin. Furthermore, studies have shown that labyrinthin is accessible to the immune system and can be targeted for immunotherapeutic interventions. The convenient location of labyrinthin on the cell surface of adenocarcinomas allows for easier accessibility and targeting by diagnostic tools and therapeutic agents. Furthermore, labyrinthin has been found to appear throughout the A549 human lung adenocarcinoma cell cycle, indicating its potential role in tumor development and progression. Additionally, labyrinthin is not modulated from the cell surface, further highlighting its stability and potential as a reliable marker for adenocarcinomas. The identification and characterization of labyrinthin as a pan-adenocarcinoma target holds significant promise for further study by clinicians and cancer researchers. Furthermore, the data provide a basis to determine to what extent labyrinthin can distinguish adenocarcinomas from other cancer types.
Challenges and future directions
While the potential of cancer neoantigens is exciting, there are challenges that need to be addressed: (1) Tumor Heterogeneity: Cancer cells within a single tumor can have different mutations, leading to the presence of multiple neoantigens. Identifying the most relevant neoantigens for each patient remains a challenge. (2) Immunosuppressive Microenvironment: The tumor microenvironment can suppress immune responses, limiting the effectiveness of neoantigen-based therapies. Strategies to overcome this immunosuppression are actively being investigated. (3) Personalization and Scalability: Developing personalized neoantigen therapies requires sophisticated sequencing and bioinformatics analysis. Ensuring these processes are efficient and scalable is essential for widespread adoption.
Conclusion
Cancer neoantigens represent a revolutionary approach to cancer treatment, leveraging the immune system’s inherent ability to recognize and target abnormal cells. As our understanding of neoantigens continues to deepen and technology advances, personalized immunotherapies based on neoantigens hold the promise of transforming cancer treatment into a more targeted, effective, and less toxic endeavour. While challenges persist, the potential benefits for patients battling cancer are immense, offering hope for a brighter future in the fight against this devastating disease.
Author contribution statement
Michael Babich & James A. Radosevich: conceived and designed the manuscript, analysed and interpreted the data, wrote the manuscript. Ankit Sharma: analysed and interpreted the data, wrote the manuscript, prepared the diagrams, performed literature survey and incorporated references
Declaration: Michael Babich and James Radosevich are principals, and Ankit Sharma is a research fellow, at LabyRx Immunological Therapeutics (USA) Limited.
References
- Esposito D, Stephen AG, Turbyville TJ, Holderfield M, editors. New weapons to penetrate the armor: Novel reagents and assays developed at the NCI RAS Initiative to enable discovery of RAS therapeutics. Seminars in cancer biology. Elsevier. 2019.
- Burrell RA, McGranahan N, Bartek J, Swanton C. The causes and consequences of genetic heterogeneity in cancer evolution. Nature. 2013; 501(7467): 338-45.
- Ostrem JM, Peters U, Sos ML, Wells JA, Shokat KM. K-Ras (G12C) inhibitors allosterically control GTP affinity and effector interactions. Nature. 2013; 503(7477): 548-51.
- Zhang J, Bu X, Wang H, Zhu Y, Geng Y, et al. Cyclin D-CDK4 kinase destabilizes PD-L1 via cullin 3-SPOP to control cancer immune surveillance. Nature. 2018; 553(7686): 91-5.
- Dagogo-Jack I, Shaw AT. Tumour heterogeneity and resistance to cancer therapies. Nature reviews Clinical oncology. 2018; 15(2): 81-94.
- Kim S-J, Lee HJ, Kim MS, Choi HJ, He J, et al. Macitentan, a dual endothelin receptor antagonist, in combination with temozolomide leads to glioblastoma regression and long-term survival in mice. Clinical Cancer Research. 2015; 21(20): 4630-41.
- Flaherty KT, Puzanov I, Kim KB, Ribas A, McArthur GA, et al. Inhibition of mutated, activated BRAF in metastatic melanoma. New England Journal of Medicine. 2010; 363(9): 809-19.
- Yaeger R, Cercek A, Chou JF, Sylvester BE, Kemeny NE, et al. BRAF mutation predicts for poor outcomes after metastasectomy in patients with metastatic colorectal cancer. Cancer. 2014; 120(15): 2316-24.
- Xing M. BRAF mutation in thyroid cancer. Endocrine-related cancer. 2005; 12(2): 245-62.
- Tiacci E, Schiavoni G, Martelli MP, Boveri E, Pacini R, et al. Constant activation of the RAF-MEK-ERK pathway as a diagnostic and therapeutic target in hairy cell leukemia. haematologica. 2013; 98(4): 635.
- Marampon F, Ciccarelli C, Zani BM. Biological rationale for targeting MEK/ERK pathways in anti-cancer therapy and to potentiate tumour responses to radiation. International journal of molecular sciences. 2019; 20(10): 2530.
- Schoor OG, Hilf NG, Weinschenk T, Trautwein CG, Walter SG, et al. Patent 2739387 Summary.
- Sampson JH, Heimberger AB, Archer GE, Aldape KD, Friedman AH, et al. Immunologic escape after prolonged progression-free survival with epidermal growth factor receptor variant III peptide vaccination in patients with newly diagnosed glioblastoma. Journal of clinical oncology. 2010; 28(31): 4722.
- O’Rourke DM, Nasrallah MP, Desai A, Melenhorst JJ, Mansfield K, et al. A single dose of peripherally infused EGFRvIII-directed CAR T cells mediates antigen loss and induces adaptive resistance in patients with recurrent glioblastoma. Science translational medicine. 2017; 9(399): eaaa0984.
- Bonavia R, Mukasa A, Narita Y, Sah DW, Vandenberg S, et al. Tumor heterogeneity is an active process maintained by a mutant EGFR-induced cytokine circuit in glioblastoma. Genes & development. 2010; 24(16): 1731-45.
- Van den Bent MJ, Vogelbaum MA, Wen PY, Macdonald DR, Chang SM. End point assessment in gliomas: Novel treatments limit usefulness of classical Macdonald’s Criteria. Journal of Clinical Oncology. 2009; 27(18): 2905.
- Wainwright DA, Chang AL, Dey M, Balyasnikova IV, Kim CK, et al. Durable therapeutic efficacy utilizing combinatorial blockade against IDO, CTLA-4, and PD-L1 in mice with brain tumors. Clinical cancer research. 2014; 20(20): 5290-301.
- Akhavan D, Alizadeh D, Wang D, Weist MR, Shepphird JK, et al. CAR T cells for brain tumors: Lessons learned and road ahead. Immunological reviews. 2019; 290(1): 60-84.
- Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nature medicine. 2013; 19(11): 1423-37.
- Xie C, Subhash VV, Datta A, Liem N, Tan SH, et al. Melanoma associated antigen (MAGE)-A3 promotes cell proliferation and chemotherapeutic drug resistance in gastric cancer. Cellular Oncology. 2016; 39: 175-86.
- Gao X, Li Q, Chen G, He H, Ma Y. MAGEA3 promotes proliferation and suppresses apoptosis in cervical cancer cells by inhibiting the KAP1/p53 signaling pathway. American Journal of Translational Research. 2020; 12(7): 3596.
- Yu QY, Wang ZW, Zhou MY, Li S-F, Liao XH. MAGE-A3 regulates tumor stemness in gastric cancer through the PI3K/AKT pathway. Aging (Albany NY). 2022; 14(23): 9579.
- Vansteenkiste J, Zielinski M, Linder A, Dahabreh J, Gonzalez EE, et al. Adjuvant MAGE-A3 immunotherapy in resected non-small-cell lung cancer: Phase II randomized study results. Journal of Clinical Oncology. 2013; 31(19): 2396-403.
- Tyagi P, Mirakhur B. MAGRIT: the largest-ever phase III lung cancer trial aims to establish a novel tumor-specific approach to therapy. Clinical lung cancer. 2009; 10(5): 371-4.
- Adams S, Robbins FM, Chen D, Wagage D, Holbeck SL, et al. HLA class I and II genotype of the NCI-60 cell lines. Journal of translational medicine. 2005; 3: 1-8.
- Atanackovic D, Altorki NK, Stockert E, Williamson B, Jungbluth AA, et al. Vaccine-induced CD4+ T cell responses to MAGE-3 protein in lung cancer patients. The Journal of Immunology. 2004; 172(5): 3289-96.
- Van Baren N, Brasseur F, Godelaine Dl, Hames Gr, Ferrant A, et al. Genes encoding tumor-specific antigens are expressed in human myeloma cells. Blood, the Journal of the American Society of Hematology. 1999; 94(4): 1156-64.
- Dreno B, Thompson JF, Smithers BM, Santinami M, Jouary T, et al. MAGE-A3 immunotherapeutic as adjuvant therapy for patients with resected, MAGE-A3-positive, stage III melanoma (DERMA): A double-blind, randomised, placebo-controlled, phase 3 trial. The Lancet Oncology. 2018; 19(7): 916-29.
- Maurer D, Butterfield LH, Vujanovic L. Melanoma vaccines: clinical status and immune endpoints. Melanoma research. 2019; 29(2): 109.
- Pan Y, Zhang Y, Chen Q, Tao X, Liu J, et al. CTAB enhances chemo-sensitivity through activation of AMPK signaling cascades in breast cancer. Frontiers in Pharmacology. 2019; 10: 843.
- Jäger E, Chen YT, Drijfhout JW, Karbach J, Ringhoffer M, et al. Simultaneous humoral and cellular immune response against cancer–testis antigen NY-ESO-1: definition of human histocompatibility leukocyte antigen (HLA)-A2-binding peptide epitopes. The Journal of experimental medicine. 1998; 187(2): 265-70.
- Odunsi K, Jungbluth AA, Stockert E, Qian F, Gnjatic S, et al. NY-ESO-1 and LAGE-1 cancer-testis antigens are potential targets for immunotherapy in epithelial ovarian cancer. Cancer research. 2003; 63(18): 6076-83.
- Wolchok JD, Weber JS, Hamid O, Lebbé C, Maio M, et al. Ipilimumab efficacy and safety in patients with advanced melanoma: A retrospective analysis of HLA subtype from four trials. Cancer Immunity. 2010; 10(1).
- Gnjatic S, Nishikawa H, Jungbluth AA, Güre AO, Ritter G, et al. NY‐ ESO‐1: Review of an immunogenic tumor antigen. Advances in cancer research. 2006; 95: 1-30.
- Robbins PF, Kassim SH, Tran TL, Crystal JS, Morgan RA, et al. A pilot trial using lymphocytes genetically engineered with an NY-ESO-1-reactive T-cell receptor: long-term follow-up and correlates with response. Clinical Cancer Research. 2015; 21(5): 1019-27.
- Ott PA, Hu Z, Keskin DB, Shukla SA, Sun J, et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature. 2017; 547(7662): 217-21.
- Slamon DJ, Leyland-Jones B, Shak S, Fuchs H, Paton V, et al. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. New England journal of medicine. 2001; 344(11): 783-92.
- Nahta R, Yu D, Hung MC, Hortobagyi GN, Esteva FJ. Mechanisms of disease: Understanding resistance to HER2-targeted therapy in human breast cancer. Nature clinical practice Oncology. 2006; 3(5): 269-80.
- Mauro C, Capone V, Cocchia R, Cademartiri F, Riccardi F, et al. Cardiovascular Side Effects of Anthracyclines and HER2 Inhibitors among Patients with Breast Cancer: A Multidisciplinary Stepwise Approach for Prevention, Early Detection, and Treatment. Journal of Clinical Medicine. 2023; 12(6): 2121.
- Baselga J, Cortés J, Kim SB, Im SA, Hegg R, et al. Pertuzumab plus trastuzumab plus docetaxel for metastatic breast cancer. New England Journal of Medicine. 2012; 366(2): 109-19.
- Levine AJ, Oren M. The first 30 years of p53: Growing ever more complex. Nature reviews cancer. 2009; 9(10): 749-58.
- Muller PA, Vousden KH. p53 mutations in cancer. Nature cell biology. 2013; 15(1): 2-8.
- Muller PA, Vousden KH. Mutant p53 in cancer: New functions and therapeutic opportunities. Cancer cell. 2014; 25(3): 304-17.
- Freed-Pastor WA, Prives C. Mutant p53: One name, many proteins. Genes & development. 2012; 26(12): 1268-86.
- Oren M, Rotter V. Introduction: p53-the first twenty years. Cellular and Molecular Life Sciences CMLS. 1999; 55: 9-11.
- Calin GA, Croce CM. Chromosomal rearrangements and microRNAs: A new cancer link with clinical implications. The Journal of clinical investigation. 2007; 117(8): 2059-66.
- Hasty P, Montagna C. Chromosomal rearrangements in cancer: detection and potential causal mechanisms. Molecular & cellular oncology. 2014; 1(1): e29904.
- Radosevich JA, Ma Y, Lee I, Salwen HR, Gould VE, et al. Monoclonal antibody 44-3A6 as a probe for a novel antigen found on human lung carcinomas with glandular differentiation. Cancer research. 1985; 45(11 Part 2): 5808-12.
- Combs S, Radosevich J, Ma Y, Lee I, Gould V, et al. Expression of the antigenic determinant recognized by the monoclonal antibody 44-3A6 on select human adenocarcinomas and normal human tissues. Tumor Biology. 1988; 9(2-3): 116-22.
- Piehl MR, Gould VE, Warren WH, Lee I, Radosevich JA, et al. Immunohistochemical identification of exocrine and neuroendocrine subsets of large cell lung carcinomas. Pathology-Research and Practice. 1988; 183(6): 675-82.
- Radosevich J, Noguchi M, Rosen S, Shimosato Y. Immunohistochemical analysis of human adenocarcinomas of the lung using the monoclonal antibody 44-3A6. Tumor Biology. 1990; 11(4): 181-8.
- Radosevich J, Combs S, Rosen S. Expression of the epitope recognized by the monoclonal antibody 44-3A6 during human fetal development. Tumor biology. 1991; 12(6): 321-9.
- Radosevich J, Siddiqui F, Rosen S, Kabat W. Cell cycle and electron microscopic evaluation of the adenocarcinoma antigen recognised by the monoclonal antibody 44-3A6. The British journal of cancer Supplement. 1991; 14: 86.
- Ordóñez NG. The value of antibodies 44-3A6, SM3, HBME-1, and thrombomodulin in differentiating epithelial pleural mesothelioma from lung adenocarcinoma: A comparative study with other commonly used antibodies. The American journal of surgical pathology. 1997; 21(12): 1399-408.
- Radosevich JA, inventor United States Patent: Cancer marker protein and peptides thereof. USA2000.
- Wands J, S De La Monte S, Deutch A, Ghanbari H, inventors. Diagnosis and treatment of malignant neoplasms. USA2001.
- Radosevich JA, Babich M Labyrinthin. The Tumor Marker Recognized By MCA 44-3A6: A Case For Pan-Tumor Markers As Targets To
Treat Cancer. Onco Targets and therapy. 2019; 12: 9351.
- Babich M, Sharma A, Li T, Radosevich JA. Labyrinthin: A distinct
pan-adenocarcinoma diagnostic and immunotherapeutic tumor
specific antigen. Heliyon. 2022; 8(3).
- Ma W, Zeng J, Montoya DJ, Toomey K, Zhou C, Chen S, et al. Labyrinthin expression is associated with poor prognosis in patients
with non-small-cell lung cancer. Cancers. 2023; 15(3): 924.
- Sharma A, Babich M, Li T, Radosevich JA. Topology and adenocarcinoma cell localization dataset on the labyrinthin diapeutic biomarker. BMC research notes. 2023; 16(1): 139.