Introduction
Despite numerous modern advances, cancer remains to be a leading cause of mortalities worldwide, estimating upto 9.7 million deaths in 2022, according recent GLOBOCAN statistics [1]. Multiple strategies tackling this challenging disease namely, surgery, chemotherapy, radiotherapy, and immunotherapy [2,3], have also resulted in therapy resistance and failure [4-7]. In this context, mounting evidences confirm a small subset of cancer stem cells (CSCs) within the tumor microenvironment (TME) that plays a crucial role in tumor initiation, metastasis, disease progression, and importantly, drug-resistance, along with immune-evasion [8-12]. A recent study from our laboratory confirms that tumor-initiating CSCs, even when present in low numbers are able to convert effector CD4+ T cells, a crucial player of anti-tumor immunity to regulatory T (Treg) cells, a pro-tumor immune fraction, by secreting TGFβ [13]. These observations substantiated the role of CSCs in immune-editing, specially involving T cells, and hence, in the possible failure of immunotherapy.
Cancer immunotherapy (CIT) potentiates the patient’s immune system to fight cancer. CIT has been documented to be effective in improving patient survival and is less toxic than conventional treatments [14]. Types of CIT techniques where evidently, T cells are found to play an active role [14,15] include Immune-Checkpoint Inhibitors (ICI), Adoptive Cell Transfer (ACT), Chimeric Antigen Receptor (CAR) T cell therapy, and cancer vaccines. However, only 20-40 percent of patients are observed to respond to immunotherapy [16]. Resistance to immunotherapy can be a consequence of dysfunction or exhaustion of T cells, as well as, the escalation of immunosuppressive, low T cell-infiltrating TME [14,17].
In this review, we discuss about different T cell-mediated immunotherapy techniques devised against cancer. Further, we highlight the probable causes underlying immunotherapy failure, wherein CSCs play a major role mainly by (1) generating Treg cells, (2) manipulating antigen presentation, and (3) increasing their own pool during therapies, all of which eventually result in immunosuppressive TME. Hence, a combinatorial application of CSC-targeting agent(s) and immunotherapy which might derive positive clinical outcomes, has also been discussed.
CIT involving T cells
Different modalities of CIT that activate the anti-tumor immune subsets, mainly, CD8+ and CD4+ T cells, are in use for cancer remission [18]. Interestingly, several studies demonstrate CD8+ T cells rely on CD4+ T cells to carry out their effector functions [19]. The activity, proliferation, and recruitment of CD8+ T cells to the tumor site are enhanced by CD4+ T cells through IL2 [20]. Therefore, effector CD4+ T cells maintains a holistic approach towards CIT. Some of the main immunotherapy avenues harnessing T cells are discussed below.
Immune-Checkpoint Inhibitors (ICI)
Immune-checkpoints, PD1 and CTLA4 are co-inhibitory receptors on the T cell surface that regulate T cell response. However, tumor cells make use of these inhibitory molecules to promote tumor tolerance and exhaustion in T cells [21]. Hence, ICIs like anti-CTLA4, anti-PD1, and anti-PDL1 can bind to these receptors, thus reinvigorating the immune response against cancer cells [21,22]. In multiple clinical trials, usage of ipilimumab (anti-CTLA4) and Pembrolizumab (anti-PD1) resulted in increased circulating CD4+ T cells and Th1 cytokines, as well as a positive outcome in patient survival [14,23]. Other clinical trials also depicted that pembrolizumab enhances overall survival if used alone, or with chemotherapy, over standard treatments [21]. Several other immune-checkpoints like, Lymphocyte Activating Gene-3 (LAG3) and T cell Immunoglobulin and ITIM domain (TIGIT) expressed on T cells, have also been the subjects of research and antibody-mediated treatment options in several cancers [24].
Adoptive cell transfer (ACT)
This method uses autologous T cells isolated from cancer patients, then identifying and isolating those with anti-tumor function, followed by expanding them ex vivo and introducing back to the patients [25]. This mechanism has been observed to be the most effective one in metastatic melanoma patients where tumor regression was found in almost 50 percent of the patients [25]. Moreover, administration of T cell- activating IL2 enhanced the effectiveness of these effector cells in mice model [26]. The combinatorial CD8+ and CD4+ T cell transfer in mouse metastasis models also showed promising outcomes [27]. In fact, owing to the lower infiltrating status of CD8+ T cells, CD4+ T cells were seen to infiltrate the tumor sites where they then stimulated tumor antigen specific-CD8+ T cells [27]. In metastatic epithelial cancer, Tran et al. [28], showed disease regression in patients when more than 95 percent of pure CD4+ T helper 1 cells were utilized.
Chimeric antigen receptor (CAR) T cell therapy
CAR-T cell therapy, another facet of T cell transfer where engineered T cells, better suited for recognizing tumor antigens and causing lymphocyte stimulation, are used. These engineered T cells present themselves as attractive subject for new generation- immunotherapy [14,29]. CARs are chimeric proteins composed of antigen binding domains, which are selected single-chain fragment variable from antibodies and one or more intracellular signaling domains of T cell receptor along with additional costimulatory domains [30]. CARs address the drawbacks of previously engineered and modified T Cell Receptor (TCR) by overcoming the need for Major Histocompatibility Complex (MHC) expression, MHC identity, and co-stimulation. This constitutes a fundamental advantage over immune evasion in cancer, as tumor cells often down-regulate MHC molecules [31]. However, CAR still requires the presence of extracellular targets on cancer cell surface [31]. CAR therapies targeting CD19 have been shown to achieve complete remission in 70 to 90 percent of patients with relapsed and refractory pre-B cell acute lymphoblastic leukemia [32], and demonstrated remarkable responses in B cell lymphomas [32]. In fact, only a small dose of CAR-T cells has been found to be effective in eradication of disease load [32]. Interestingly, CD4+ T cells were as effective as CD8+ T cells towards implementing cytotoxicity, and showed lower exhaustion compared to CD8+ T cells in CD19 targeted-CAR therapy in mice models for lymphoblastic leukemia [14,32].
Therapeutic cancer vaccination
Much like the vaccination mechanisms eliciting immune responses, cancer antigens or personalized neoantigens, after immunization, are processed by Antigen Presenting Cells (APCs) and express them on MHC I/II complexes, which in turn, prime and activate T cell response. This further propagates and leads to T cell infiltration to tumor site [33]. Peptide vaccine against HPV-16 positive intraepithelial vulvar neoplasia resulted in high IFNγ responses by CD4+ and CD8+ T cells [34]. Moreover, CD4+ T cells were observed to have higher IFNγ expression in patients demonstrating complete response [34]. A long lasting immune response with robust CD4+ T cell levels was observed in a phase I/II clinical trials with prostate cancer patients when treated with peptide vaccine targeting RhoC [35].
Limitations and failure of cancer T cell immunotherapy
Although these varied immunotherapy strategies have proven effective in multiple cancers and generated durable response, they have often resulted to therapy- resistance and failure [5,36]. For example, T cell dysfunction as a consequence of chronic antigen stimulation causes loss of its effector and anti-tumor functionalities [14]. On the other hand, an immunosuppressive tumor milieu, characterized by high levels of cancer-aiding factors like TGFβ, VEGF, and IL8, metabolites like, Indoleamine 2, 3-Dioxygenase (IDO), low antigenicity, lack of immune ‘call signal’ chemokines, dysregulated signaling pathways, and most importantly, the presence of tumor-promoting immune subsets, results in exhaustion and lower infiltration of active effector T cells [5]. In this context, the infiltration of Tregs and conversion of anti-tumor T cells to Treg cells play quite a significant role in defining immunotherapy resistance [17,37]. Tumor-promoting Tregs play a crucial role in suppressing the body’s immune function against tumors, while also aiding in Epithelial-Mesenchymal Transition (EMT), angiogenesis, and invasion [13,37]. Presence of increased Tregs and their infiltration at the tumor site leads to a negative clinical prognosis [38].
As directed by our previous report and other concurrent findings [13,39], the marked role of CSCs towards shifting the anti-tumor T cell population to pro-tumor Tregs is well understood. Consequently, immunotherapy strategies discussed above, aimed at enhancing effector T cell numbers and their activity, although portray an interesting prospect, might be facing the hurdle of therapy resistance mainly conferred by the interplay of Treg cells and CSCs [13,37].
Cancer stem cells
Other than tumor cells, the TME consists of immune cells, the extracellular matrix, exosomes, cytokines, and cell-secreted cytokines [40]. CSCs are a small subset of tumor cells that have been identified as the primary cause of cancer initiation, along with treatment failures, cancer recurrence, tumor growth, invasion, and metastasis, as supported by increasing evidences [8,41]. These tiny sub-population of cancer cells can even escape the stress signals such as radiotherapy, chemotherapy drugs by utilizing different modalities to hide in the TME [8,41]. Most importantly, during the early stages of tumor development, tumor initiating-CSCs must avoid immune- surveillance and immune-editing to survive in the TME for propagating and populating the entire tumor mass [8,43,44]. CSCs accomplish this difficult task by evading identification and restricting anti-tumor actions by the immune subsets, or by regulating several TME compartments, including immune and non-immune ones, in their favor [8,41]. These immune-evading properties of CSCs support their possible seminal roles in CIT failure.
Role of CSCs in T cell-immunotherapy failure
Effector T cells primed and induced by CIT, fail to furnish their ‘original’ anti-tumor function, mainly due to the following reasons.
Treg generation
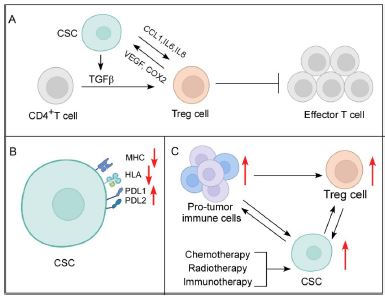
Our previous study reported a strong correlation between CSCs with Tregs in tumor [13]. In fact, our report showed that very low number of CSCs could successfully convert infiltrating CD4+ T lymphocytes to Treg cells in a contact-independent manner. Moreover, these CSCs not only escaped chemotherapy, but also generated more Treg cells. Our exploration further reported that CSCs generated Treg cells from effector T cells by secreting immune-suppressive, Treg-polarizing cytokine TGFβ, which was further exacerbated by chemotherapy [13]. Similarly, other studies corroborated that under hypoxic effects, glioblastoma CSCs inhibited T cell functions, induced FOXP3+ Tregs and generated immunosuppressive TME [43]. Additionally, SOX-expressing CSCs release CCL1, which attracts Tregs to the TME, the Tregs in turn produce TGFβ and IL17, augmenting self-renewal, cancer stemness, and EMT towards tumor development and invasion [44]. Reportedly, STAT3 signaling protects gastric CSCs from T cell-mediated death and differentiates uncommitted CD4+T cells into Tregs [45]. CSCs also influence the Th17/Treg balance by altering the levels of IL6, IL8, and CCL5 in the TME [46]. TGFβ further enables CD80 ligand-expressing CSCs to interact with Tregs and CTLA4, resulting in diminished cytotoxic T cell activity and immunotherapy resistance [47]. Therefore, these findings strongly nudge towards the impact of CSCs in promotion of Tregs.
Conversely, Tregs release VEGF, which promotes angiogenesis and thereby aids in CSC survival, stemness modulation, and their self-renewal in hypoxic condition [48]. Treg-derived cyclooxygenase 2 (COX2) inhibits effector T cells via PEG-E2- dependent mechanism [47]. The release of TGFβ, IL10, and IL35 by Tregs hinders T cell proliferation and effector functions, including IFNγ generation [13,49] and granzyme and perforin-mediated killing, thus, contributing to immunotherapy failure [50] (Figure 1). There, therefore, exists a deadly liaison between CSCs and Treg cells in TME that might affect the fate of CIT.
Antigen presentation
The immune system targets cancer cells through antigen processing and presentation [51]. APCs detect cancer cell-associated antigens and display them on their surfaces in order to be recognized by T cells [51]. Unfortunately, CSCs avoid the immune system by down-regulating MHC expression or inhibiting the expression of antigens identified by immune cells [51,52]. Head and neck cancer CD44+ CSCs have been documented to downregulate Human Leukocyte Antigen-A2 (HLA-A2), HLA class II, and TAP2 expression [53]. Whereas, melanoma CSCs apart from expressing low MHCI and MHCII, circumvent tumor immune surveillance by inhibiting transcription of melanoma-associated antigens as well [52].
Also, PDL1 and PDL2, which are overexpressed in CSCs, inhibit T cell activity and allow immunological escape [41,52] (Figure 1). In addition, interaction of PDL1 and PDL2 with PD1+ T cells releases IL10 from tumor cells, which then suppresses effector T cells [54], thereby, dampening the functions of T cells in multiple ways. By activating PI3K/AKT and mTOR signaling pathways, PDL1 also maintains CSC pluripotency [55]. Moreover, CSCs show heightened expression of CD47, which interferes with the phagocytic activity of tumor-associated macrophages (TAMs), hence, limit their role as APCs, and consequently, T cell priming and activation [8].
Cancer therapies augment CSC population and further add to immunosuppressive TME
Multiple treatment regimens, such as, chemotherapy, radiotherapy, and even, immunotherapy enhance CSC nature and its pool [10,58-60]. Previous report from our laboratory demonstrated that chemotherapy not only failed to induce apoptosis in CSCs, it also enhanced the CSC pool via de-differentiation of Non-Stem Cancer Cells (NSCCs)[10]. CSCs also showed radio-resistance while radiotherapy promoted EMT and generation of CSCs [56].
In fact, immunotherapy has also been observed to increase CSC nature in breast cancer [58]. Since CSCs and Tregs have a strong association, increase in CSC population during different treatment modalities, invariably amounts to increase in Treg numbers [13]. Other components of the TME, such as TAMs, being promoted by CSCs, also end up aiding Treg cells [8,59]. Myeloid- Derived Suppressor Cells (MDSCs) are recruited by TGFβ-activated CD133+ CSCs in melanoma [47]. Incidentally, MDSCs are reported to stimulate Tregs and TAMs [60]. Again, Tregs, TAMs, and MDSCs promote CSC status in tumor [41,47], thereby, maintaining an overall crosstalk, leading to immunosuppressive TME and impaired infiltration of anti-tumor T cells [61] (Figure 1).
Therefore, challenges for immunotherapy measures like, ACT and vaccines, which aim at increasing active T cells, eventually may end up being ineffective due to the T cell to Treg cell converting potential of CSCs [13]. As discussed above, additional therapies may worsen the situation, accounting for enhanced levels of CSCs and pro- tumor subsets, resulting in ‘cold’ tumor status and immunotherapy failure [13,41,61]. Hence, it was not unexpected when poor efficacy, clinical outcomes, and exhaustion of T cells were seen as limitations of ACT and cancer vaccines [33,62]. Notably, TGFβ- responsive CSCs were observed to be ACT resistant [63]. Other than high immune- checkpoint expression, CSCs demonstrate intrinsic and extrinsic resistance factors, which are hindrance to ICI therapy [36]. Supporting this notion, a study revealed a positive correlation of CSC marker with genes associated with ICI-immunotherapy resistance [64]. Additionally, similar to ACT, CAR-T faces challenges owing to low T- cell infiltration and low cancer antigen expression [52,65] (Figure 2).
Possible remedy
In essence, combinatorial therapy involving sensitization and elimination of CSCs followed by conventional therapies might sustain an anti-tumor TME. However, therapeutic stress induces NSCCs to de-differentiate into CSCs [10], and again establish an immunosuppressive environment. We found that pre-treating CSCs with aspirin sensitizes them to chemo-treatment [10]. By down-regulating WNT pathway with paclitaxel, CSC content and tumor development were considerably decreased [66,67]. Likewise, Metformin has been reported to alter epigenetic landscape of CSCs, rendering them vulnerable to chemotherapy [68].
Monoclonal antibodies (mAbs) have recently been proved effective and garnered acceptance in cancer therapy. Multiple clinical trials have employed mAbs to target CSC-associated biomarkers. For example, CD20, a common CSC marker was targeted by rituximab which effected favourable response rate in lymphoma [69]. CSC marker, EpCAM was used as a target by employing Adecatumumab, that demonstrated remarkable therapeutic potential in hormone-resistant breast cancer patients [70].
Recently, CAR-T cells targeting CSC-specific antigens have been tested in cancer models. CAR-T cells against EpCAM antigens have successfully eliminated CSCs in PC3M and PC3 prostate cancer cell line models [71]. CSCs were selectively killed by adoptively transplanted cytotoxic T cells specific for ASB4 antigen in colon cancer [72]. Adoptively transferring CAR natural killer (NK) cells have also shown to eliminate CSCs with high efficiency [47].
Dendritic Cell (DC)-based vaccinations are found to be effective against CSCs. DCs pulsed with cancer cell lines or CSC-lysates were applied as vaccines to test therapeutic effectiveness. In the malignant melanoma model, CSC-lysate-pulsed DCs decreased tumor development and prolonged life in vaccinated mice by inducing IFNγ and IL4 production [73,74]. DCs charged with Panc-1 CSC-lysates stimulated INFγ and IL2 production, leading to lymphocyte infiltration in pancreatic cancer [75]. DCs with NANOG peptides generated highly specific anti-tumor T cell responses against ovarian cancer CSCs [76].
However, detail studies are required to authentically identify the possible agents/molecules/processes that will successfully kill/sensitize CSCs, thereby, will pave the way for successful immunotherapy in combinatorial treatment.
Concluding remarks
Above discussion highlights different T cell based-immunotherapy modalities available, as well as, their limitations. We have further elaborated that CSCs play a crucial part in immunotherapy-resistance by generating or recruiting Tregs, limiting effector T cell activity, presenting lower antigens, expressing high immune check-points and creating a cumulatively interacting immunosuppressive TME, thereby impairing anti- tumor T cell infiltration. To overcome such obstacles, potent therapies targeting CSC-associated antigens, alone or in conjunction with CSC-supportive TME, must be considered to be used prior to immunotherapy. As CSCs share tumor-initiating and immune evasion features, immunotherapies tailored against CSC-specific neoantigens and stemness transcription factors, are of utmost need of present day. However, ample research involving CSC-targeted CAR-T/ DC/NK cell cancer therapies are required to ensure clinical safety and efficacy, assuring their use in future interventions for ensuring overall success of CIT.
Declarations
Conflicts of interest: The authors have no competing interests to declare that are relevant to the content of this article.
Funding: This study was supported by the grants received from: ICMR Emeritus Scientist Scheme (TD; 74/1/2020-Pers).
Author contributions: Conceptualization, supervision, review, and editing: Tanya Das; Writing-original draft: Udit Basak, Sourio Chakraborty; Visualization: Sumon Mukherjee.
References
- Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians. 2024; 74(3): 229-63.
- Debela DT, Muzazu SG, Heraro KD, Ndalama MT, Mesele BW, et al. New approaches and procedures for cancer treatment: Current perspectives. SAGE Open Med. 2021; 9: 20503121211034366.
- Treatment for Cancer - NCI. 2015. https: //www.cancer.gov/about-cancer/treatment
- Vasan N, Baselga J, Hyman DM. A view on drug resistance in cancer. Nature. 2019; 575(7782): 299-309.
- Błach J, Wojas-Krawczyk K, Nicoś M, Krawczyk P. Failure of Immunotherapy- The Molecular and Immunological Origin of Immunotherapy Resistance in Lung Cancer. Int J Mol Sci. 2021; 22(16): 9030.
- Rukstalis DB. Treatment Options after Failure of Radiation Therapy-A Review. Rev Urol. 2002; 4(Suppl 2): S12-7.
- Mirzayans R, Murray D. What Are the Reasons for Continuing Failures in Cancer Therapy? Are Misleading/Inappropriate Preclinical Assays to Be Blamed? Might Some Modern Therapies Cause More Harm than Benefit? Int J Mol Sci. 2022; 23(21): 13217.
- Basak U, Sarkar T, Mukherjee S, Chakraborty S, Dutta A, et al. Tumor- associated macrophages: an effective player of the tumor microenvironment. Front Immunol. 2023; 14: 1295257.
- Mukherjee S, Manna A, Bhattacharjee P, Mazumdar M, Saha S, et al. Non-migratory tumorigenic intrinsic cancer stem cells ensure breast cancer metastasis by generation of CXCR4+ migrating cancer stem cells. Oncogene. 2016; 35(37): 4937-48.
- Saha S, Mukherjee S, Khan P, Kajal K, Mazumdar M, et al. Aspirin Suppresses the Acquisition of Chemoresistance in Breast Cancer by Disrupting an NFκB-IL6 Signaling Axis Responsible for the Generation of Cancer Stem Cells. Cancer Res. 2016; 76(7): 2000-12.
- Banerjee S, Mukherjee S, Bhattacharya A, Basak U, Chakraborty S, et al. Pyridoxine enhances chemo-responsiveness of breast cancer stem cells via redox reconditioning. Free Radic Biol Med. 2020; 152: 152-65.
- Bhattacharya A, Mukherjee S, Khan P, Banerjee S, Dutta A, et al. SMAR1 repression by pluripotency factors and consequent chemoresistance in breast cancer stem-like cells is reversed by aspirin. Sci Signal. 2020; 13(654): eaay6077.
- Mukherjee S, Chakraborty S, Basak U, Pati S, Dutta A, et al. Breast cancer stem cells generate immune-suppressive T regulatory cells by secreting TGFβ to evade immune-elimination. Discov Onc. 2023; 14(1): 220.
- Ahmed H, Mahmud AR, Siddiquee MohdFR, Shahriar A, Biswas P, et al. Role of T cells in cancer immunotherapy: Opportunities and challenges. Cancer Pathogenesis and Therapy. 2023; 1(2): 116-26.
- Immunotherapy for Cancer - NCI. 2015. https: //www.cancer.gov/about-cancer/treatment/types/immunotherapy
- Decoding the signs of response to cancer immunotherapy. 2024. https: //www.nature.com/articles/d42473-019-00064-0
- Tanaka A, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017; 27(1): 109-18.
- Richardson JR, Schöllhorn A, Gouttefangeas C, Schuhmacher J. CD4+ T Cells: Multitasking Cells in the Duty of Cancer Immunotherapy. Cancers (Basel). 2021; 13(4): 596.
- Keene JA, Forman J. Helper activity is required for the in vivo generation of cytotoxic T lymphocytes. Journal of Experimental Medicine. 1982; 155(3): 768-82.
- Bos R, Sherman LA. CD4+ T-Cell Help in the Tumor Milieu is required for Recruitment and Cytolytic Function of CD8+ T Lymphocytes. Cancer Research. 2010; 70(21): 8368-77.
- Shiravand Y, Khodadadi F, Kashani SMA, Hosseini-Fard SR, Hosseini S, et al. Immune Checkpoint Inhibitors in Cancer Therapy. Curr Oncol. 2022; 29(5): 3044-60.
- Marei HE, Hasan A, Pozzoli G, Cenciarelli C. Cancer immunotherapy with immune checkpoint inhibitors (ICIs): potential, mechanisms of resistance, and strategies for reinvigorating T cell responsiveness when resistance is acquired. Cancer Cell Int. 2023; 23: 64.
- Martens A, Wistuba-Hamprecht K, Yuan J, Postow MA, Wong P, et al. Increases in Absolute Lymphocytes and Circulating CD4+ and CD8+ T Cells Are Associated with Positive Clinical Outcome of Melanoma Patients Treated with Ipilimumab. Clinical Cancer Research. 2016; 22(19): 4848-58.
- Wang DR, Wu XL, Sun YL. Therapeutic targets and biomarkers of tumor immunotherapy: Response versus non-response. Sig Transduct Target Ther. 2022; 7(1): 1-27.
- Rosenberg SA, Restifo NP, Yang JC, Morgan RA, Dudley ME. Adoptive cell transfer: a clinical path to effective cancer immunotherapy. Nat Rev Cancer. 2008; 8(4): 299-308.
- Donohue JH, Rosenstein M, Chang AE, Lode MT, Robb RJ, et al. The systemic administration of purified interleukin 2 enhances the ability of sensitized murine lymphocytes to cure a disseminated syngeneic lymphoma. J Immunol. 1984; 132(4): 2123-8.
- Wang LX, Shu S, Disis ML, Plaud GE. Adoptive transfer of tumor-primed, in vitro- activated, CD4+ T effector cells (TEs) combined with CD8+ TEs provides intratumoral TE proliferation and synergistic antitumor response. Blood. 2007; 109(11): 4865-76.
- Cancer Immunotherapy Based on Mutation-Specific CD4+ T Cells in a Patient with Epithelial Cancer. 2024. https: //www.science.org/doi/10.1126/science.1251102
- S Bridgeman J, E Hawkins R, A Hombach A, Abken H, E Gilham D. Building Better Chimeric Antigen Receptors for Adoptive T Cell Therapy. Current Gene Therapy. 2010; 10(2): 77-90.
- Miliotou AN, Papadopoulou LC. CAR T-cell Therapy: A New Era in Cancer Immunotherapy. Current Pharmaceutical Biotechnology. 2018; 19(1): 5-18.
- June CH, O’Connor RS, Kawalekar OU, Ghassemi S, Milone MC. CAR T cell immunotherapy for human cancer. Science. 2018; 359(6382): 1361-5.
- TCR engagement negatively affects CD8 but not CD4 CAR T cell expansion and leukemic clearance. 2024. https: //www.science.org/doi/10.1126/scitranslmed.aag1209
- Fan T, Zhang M, Yang J, Zhu Z, Cao W, et al. Therapeutic cancer vaccines: advancements, challenges, and prospects. Sig Transduct Target Ther. 2023; 8(1): 1- 23.
- Kenter Gemma G, Welters Marij JP, Valentijn A Rob PM, Lowik Margriet JG, Berends-van der Meer Dorien MA, et al. Vaccination against HPV-16 Oncoproteins for Vulvar Intraepithelial Neoplasia. New England Journal of Medicine. 2009; 361(19): 1838-47.
- Schuhmacher J, Heidu S, Balchen T, Richardson JR, Schmeld C, et al. Vaccination against RhoC induces long-lasting immune responses in patients with prostate cancer: results from a phase I/II clinical trial. J Immunother Cancer. 2020; 8(2): e001157.
- Bai R, Chen N, Li L, Du N, Bai L, et al. Mechanisms of Cancer Resistance to Immunotherapy. Frontiers in Oncology. 2020; 10. https: //www.frontiersin.org/articles/10.3389/fonc.2020.01290
- Togashi Y, Shitara K, Nishikawa H. Regulatory T cells in cancer immunosuppression - implications for anticancer therapy. Nat Rev Clin Oncol. 2019; 16(6): 356-71.
- Tanaka A, Sakaguchi S. Targeting Treg cells in cancer immunotherapy. Eur J Immunol. 2019; 49(8): 1140-6.
- Silver DJ, Sinyuk M, Vogelbaum MA, Ahluwalia MS, Lathia JD. The intersection of cancer, cancer stem cells, and the immune system: therapeutic opportunities. Neuro-Oncology. 2016; 18(2): 153-9.
- Sarkar T, Dhar S, Sa G. Tumor-infiltrating T-regulatory cells adapt to altered metabolism to promote tumor-immune escape. Curr Res Immunol. 2021; 2: 132-41.
- Chakraborty S, Mukherjee S, Basak U, Pati S, Dutta A, et al. Immune evasion by cancer stem cells ensures tumor initiation and failure of immunotherapy. Explor Immunol. 2023; 3(4): 384-405.
- Adam JK, Odhav B, Bhoola KD. Immune responses in cancer. Pharmacol Ther. 2003; 99(1): 113-32.
- Wei J, Wu A, Kong LY, Wang Y, Fuller G, et al. Hypoxia Potentiates Glioma- Mediated Immunosuppression. PLOS ONE. 2011; 6(1): e16195.
- Xu Y, Dong X, Qi P, Ye Y, Shen W, et al. Sox2 Communicates with Tregs through CCL1 to Promote the Stemness Property of Breast Cancer Cells. Stem Cells. 2017; 35(12): 2351-65.
- Rezalotfi A, Ahmadian E, Aazami H, Solgi G, Ebrahimi M. Gastric Cancer Stem Cells Effect on Th17/Treg Balance; A Bench to Beside Perspective. Front Oncol. 2019; 9: 226.
- Yu X, Li H, Ren X. Interaction between regulatory T cells and cancer stem cells. International Journal of Cancer. 2012; 131(7): 1491-8.
- Dianat-Moghadam H, Mahari A, Salahlou R, Khalili M, Azizi M, et al. Immune evader cancer stem cells direct the perspective approaches to cancer immunotherapy. Stem Cell Research & Therapy. 2022; 13(1): 150.
- Beck B, Driessens G, Goossens S, Youssef KK, Kuchnio A, et al. A vascular niche and a VEGF-Nrp1 loop regulate the initiation and stemness of skin tumours. Nature. 2011; 478(7369): 399-403.
- Sawant DV, Yano H, Chikina M, Zhang Q, Liao M, et al. Adaptive plasticity of IL-10+ and IL-35+ Treg cells cooperatively promotes tumor T cell exhaustion. Nat Immunol. 2019; 20(6): 724-35.
- Cao X, Cai SF, Fehniger TA, Song J, Collins LI, et al. Granzyme B and Perforin Are Important for Regulatory T Cell-Mediated Suppression of Tumor Clearance. Immunity. 2007; 27(4): 635-46.
- Kallingal A, Olszewski M, Maciejewska N, Brankiewicz W, Baginski M. Cancer immune escape: the role of antigen presentation machinery. J Cancer Res Clin Oncol. 2023; 149(10): 8131-41.
- Tsuchiya H, Shiota G. Immune evasion by cancer stem cells. Regen Ther. 2021; 17: 20-33.
- Chikamatsu K, Takahashi G, Sakakura K, Ferrone S, Masuyama K. Immunoregulatory Properties Of Cd44+ Cancer Stem-Like Cells In Squamous Cell Carcinoma Of The Head And Neck. Head Neck. 2011; 33(2): 208-15.
- Christiansson L, Lindqvist C, Tötterman TH, Simonsson B, Olsson-Strömberg U, et al. Soluble IL2R (CD25), IL10 and PDL1 May Control T Cell Activation in Chronic Myeloid Leukemia. Blood. 2009; 114(22): 4252.
- Makena MR, Ranjan A, Thirumala V, Reddy AP. Cancer stem cells: Road to therapeutic resistance and strategies to overcome resistance. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 2020; 1866(4): 165339.
- Liu Y, Yang M, Luo J, Zhou H. Radiotherapy targeting cancer stem cells “awakens” them to induce tumour relapse and metastasis in oral cancer. Int J Oral Sci. 2020; 12(1): 1-12.
- Gupta G, Merhej G, Saravanan S, Chen H. Cancer resistance to immunotherapy: What is the role of cancer stem cells? Cancer Drug Resist. 2022; 5(4): 981-94.
- Roarty K. Unlocking the secrets of cancer stem cells: Immune checkpoint inhibitors face their formidable foes. Cell Stem Cell. 2023; 30(6): 743-4.
- Xiang X, Wang J, Lu D, Xu X. Targeting tumor-associated macrophages to synergize tumor immunotherapy. Sig Transduct Target Ther. 2021; 6(1): 1-12.
- Li K, Shi H, Zhang B, Ou X, Ma Q, et al. Myeloid-derived suppressor cells as immunosuppressive regulators and therapeutic targets in cancer. Sig Transduct Target Ther. 2021; 6(1): 1-25.
- Yang W, Liu S, Mao M, Gong Y, Li X, et al. T-cell infiltration and its regulatory mechanisms in cancers: Insights at single-cell resolution. Journal of Experimental & Clinical Cancer Research. 2024; 43(1): 38.
- Kalos M, June CH. Adoptive T cell Transfer for Cancer Immunotherapy in the Era of Synthetic Biology. Immunity. 2013; 39(1): 10. 1016/j.immuni.2013.07.002.
- Miao Y, Yang H, Levorse J, Yuan S, Polak L, et al. Adaptive Immune Resistance Emerges from Tumor-Initiating Stem Cells. Cell. 2019; 177(5): 1172- 1186.e14.
- Yu W, Liu F, Lei Q, Wu P, Yang L, et al. Identification of Key Pathways and Genes Related to Immunotherapy Resistance of LUAD Based on WGCNA Analysis. Front Oncol. 2022; 11: 814014.
- Sterner RC, Sterner RM. CAR-T cell therapy: Current limitations and potential strategies. Blood Cancer J. 2021; 11(4): 1-11.
- Allavena P, Digifico E, Belgiovine C. Macrophages and cancer stem cells: A malevolent alliance. Molecular Medicine. 2021; 27(1): 121.
- Fischer MM, Cancilla B, Yeung VP, Cattaruzza F, Chartier C, et al. WNT antagonists exhibit unique combinatorial antitumor activity with taxanes by potentiating mitotic cell death. Sci Adv. 2017; 3(6): e1700090.
- Brown JR, Chan DK, Shank JJ, Griffith KA, Fan H, et al. Phase II clinical trial of metformin as a cancer stem cell-targeting agent in ovarian cancer. JCI Insight. 2020; 5(11): e133247, 133247.
- Zhou H, Tan L, Liu B, Guan XY. Cancer stem cells: Recent insights and therapies. Biochemical Pharmacology. 2023; 209: 115441.
- Schmidt M, Scheulen ME, Dittrich C, Obrist P, Marschner N, et al. An open-label, randomized phase II study of adecatumumab, a fully human anti- EpCAM antibody, as monotherapy in patients with metastatic breast cancer. Annals of Oncology. 2010; 21(2): 275-82.
- Deng Z, Wu Y, Ma W, Zhang S, Zhang YQ. Adoptive T-cell therapy of prostate cancer targeting the cancer stem cell antigen EpCAM. BMC Immunol. 2015; 16(1): 1.
- Miyamoto S, Kochin V, Kanaseki T, Hongo A, Tokita S, et al. The Antigen ASB4 on Cancer Stem Cells Serves as a Target for CTL Immunotherapy of Colorectal Cancer. Cancer Immunol Res. 2018; 6(3): 358-69.
- Dashti A, Ebrahimi M, Hadjati J, Memarnejadian A, Moazzeni SM. Dendritic cell based immunotherapy using tumor stem cells mediates potent antitumor immune responses. Cancer Lett. 2016; 374(1): 175-85.
- Badrinath N, Yoo SY. Recent Advances in Cancer Stem Cell-Targeted Immunotherapy. Cancers (Basel). 2019; 11(3): 310.
- Yin T, Shi P, Gou S, Shen Q, Wang C. Dendritic cells loaded with pancreatic Cancer Stem Cells (CSCs) lysates induce antitumor immune killing effect in vitro. PLoS One. 2014; 9(12): e114581.
- Wefers C, Schreibelt G, Massuger LFAG, de Vries IJM, Torensma R. Immune Curbing of Cancer Stem Cells by CTLs Directed to NANOG. Front Immunol. 2018; 9: 1412.