Introduction
Boron Neutron Capture Therapy Capture Therapy (BNCT) is a chemo radiation therapy. By administering drugs that contains 10-B atom, that tend to accumulate in cancer cells, irradiation of the cancer affecting area with thermal neutron beams, thermal neutron is captured and particle ion beams of the helium nucleus of helium and 7Li, with a short path range of 4-5 μm and 9-10 μm, respectively, are generated. The diameter of cancer cells is mainly several tens of micrometers, so damage such as double-stranded DNA breaks that are difficult to repair to selectively kill cancer cells. When cellular responses were analyzed, HMGB1 protein was found to be released extracellularly from cancer cells and in xenograft mouse model at early period [1].
HMGB1 is known as a chromatin-binding protein and expressed widely in the cells [2]. HMGB1 is known to be present in nucleus and has DNA binding activity and is involved in transcriptional regulation [3]. The increase of HMGB1 expression level was reported in cancer cells and its inflammatory role belonging the group of Damage-Associated Molecular Patterns (DAMPs) is reported [4-6]. Receptor for Advanced Glycation End-Product (RAGE) and Toll-Like Receptor (TLR) are known as relation to HMGB1 in terms of activation of proinflammatory ligands [7-9].
HMGB1 is involved in the DNA damage response and cell death [10-12]. Apoptotic cell death, autophagic cell death and necrotic cell death cause HMGB1 extracellular release from various types of the cells [5,13,14]. It is considered that extracellular HMGB1 release promotes inflammation reaction, sepsis, and autoimmune diseases [7]. In this study, we focused on the states and dynamic changes of HMGB1 levels at mRNA in the tumor cells after Boron Neutron Capture Therapy (BNCT) using thermal neutron beam irradiation with 10B- Boronophenylalanine (BPA) and discussed a role during cellular response to BNCT.
Materials and methods
Cell culture
Human melanoma A375 cell line and human squamous cell carcinoma SAS cell line were cultured in DMEM medium containing 10% Fetal Bovine Serum (FBS), 1% Penicillin and Streptomycin (PS) and DMEM/Ham’s F12 medium containing 10% FBS and 1% PS, respectively. Both cells were cultured in the humidified CO2 incubator at 37 °C. 10B-Enriched (>98%) BPA was purchased from Katchem spol. s.r.o. (Praha, Czech Republic). BPA fructose complex was prepared and added to the cell culture as described elsewhere [15-17].
Neutron beam irradiation and BNCT experiment
The neutron beam irradiations were operated at Kyoto University Research Reactor (KUR). Before the irradiation, cells were suspended and diluted to 1 x 106 cells/ml. At least 1 hour before the irradiation, BPA fructose complex at the boron concentration of 25 ppm [10B] or mock-control was added, respectively. The irradiation doses for BNCT were chosen based on previous reports [15] and simulated the dose to be 0, 4 and 24 Gy-Eq for BPA administrated cells. γ-Ray irradiation with 137Cs source was carried out using Gamma Cell 40 Exactor (Best Theratronics, Canada) at National Cancer Center Research Institute or Nagasaki University (PS-3100SE, Pony Industry, Japan). The dose rate was approximately 100 cGy/min. Neutron beam irradiation and γ-irradiation were was operated at suspension state.
The quantitative RT-PCR
RNA was extracted using Isogen (Nippon Gene) and reverse transcription was processed using High Capacity Reverse Transcription Kit (Thermo Fisher Scientific). The quantitative RT-PCR (qRT-PCR) analysis was performed using SYBR Green using the CFX96 Real-Time System (Bio-Rad). The normalization was carried out using GUSB mRNA. The primer pairs used are listed in Appendix Table 1.
Western blot analysis and two dimensional-polyacrylamide gel electrophoresis analysis
Protein fractions from the cells were extracted using Isogen (Dojin) as recommended by the manufacturer. Protein concentration was determined using Protein Assay Solution (Bio- Rad) and SDS polyacrylamide gel electrophoresis was carried out as previously described [10]. The protein samples were separated also by 2D gel electrophoresis and the gel images were analyzed. In gel tryptic digesting and matrix assisted laser desorption/ionization time of flight mass spectrometry were performed as described previously [16]. The used antibodies and reaction conditions were listed in Appendix Table 2.
Statistical analysis
Statistical analysis was carried out by Student’s t-tests using JMP (SAS Institute Inc. Cary, NC, USA).
Table 1: Realtime PCR primers.
| Gene |
Forward |
Human |
Reverse |
| GUSB |
GCCTGCGTCCCACCTAGAAT |
|
ACATACGGAGCCCCCTTGTC |
| HMGB1 |
ACTGGGCGACTCTGTGCCTC |
|
AGACATGGTCTTCCACCTCTCTGA |
Table 2: Antibodies used for western blot analysis and immunohistochemistry.
| Antibody |
Host |
Species reactivity |
Company |
Cat# |
| Anti-HMGB1 |
Rabbit/polyclonal |
Human, Mouse, Rat |
Abcam |
ab104774 |
Results
The analysis of changes in HMGB1 mRNA level after neutron beam irradiation with or without BPA and γ-irradiation in A375 and SAS cells.
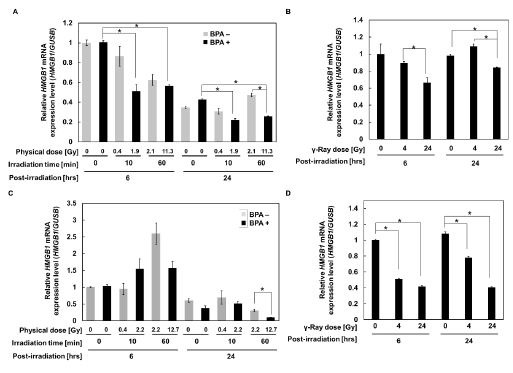
To examine whether the increase of HMGB1 release is associated with the changes in the mRNA expression level, we examined the HMGB1 mRNA levels in A375 and SAS cells after neutron beam irradiation with BPA. The real-time PCR analysis showed a decrease in the HMGB1 mRNA at 6 and 24 hrs after 60 min neutron beam irradiation in the presence of BPA compared with no irradiation control in SAS cells (Figure 1A).
After γ-irradiation, the mRNA level showed a slight dose-dependent decrease but the difference was not marked both at 6 and 24 hrs in SAS cells (Figure 1B). The HMGB1 mRNA level in A375 cells showed the lowest level 24 hrs after 60 min neutron beam irradiation in presence of BPA (Figure 1C), whereas the level decreased to around half both 6 and 24 hrs after 24 Gy γ- irradiation (Figure 1D). These results suggest that extracellular release of HMGB1 from A375 and SAS cells after BNCT or γ-irradiation occurred at rather lowered conditions of HMGB1 mRNA. Compared to SAS cells, the basal level of HMGB1 mRNA in A375 is approximately 4-fold lower. Therefore, it may be reasonable that the extracellular level of HMGB1 was not detected in A375 as we reported [1].
Status of HMGB1 protein in SAS cells after neutron beam irradiation with BPA
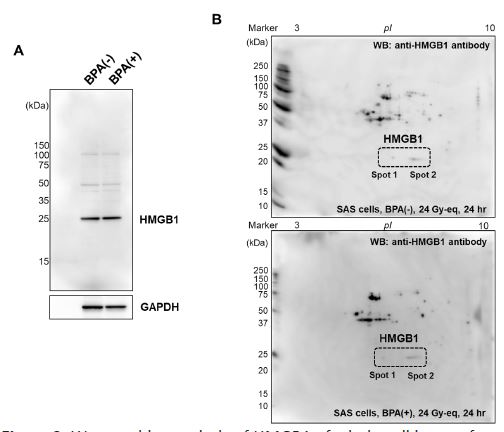
Because SAS cells showed tenfold higher level of HMGB1 release after neutron beam irradiation with BPA compared with A375 cells, we analyzed whether HMGB1 protein shows changes after neutron beam irradiation with BPA in SAS cells. HMGB1 protein has acidic residues in the C-terminal region, which has been reported to be modified with polyADP- ribosylation, leading to the release from the cells [18]. SAS cells were irradiated with neutron beam in the absence and presence of BPA (60 min neutron beam irradiation) and 24 hrs after the irradiation, the cell lysates were subjected to western blotting as shown in Figure 2A. There was no difference in HMGB1 level between SAS cells irradiated in BNCT condition with or in the absence of BPA addition. Two-dimensional gel electrophoresis was further carried out and western blotting using HMGB1 antibody was performed. As shown in Figure 2B, HMGB1 was observed as at neutral and a slightly acidic pIs, however, their amounts and ratio did not change 24 hrs after neutron beam irradiation with BPA. These results suggested that acidic modification, such as polyADP-ribosylation, was not enhanced in this condition in SAS cells after BNCT irradiation condition.
Discussion
In this study, we demonstrated that HMGB1 mRNA is rapidly decreased after BNCT of therapeutic doses in both oral cancer SAS cells and melanoma A375 cells as early period of 6-24 hrs post irradiation. In contrast, γ-irradiation of equivalent doses caused less levels of mRNA decrease in both SAS cells and A375 cells. On the other hand, at 24 hrs after BNCT, we did not observe clear changes in the property of HMGB1, suggesting protein modification on HMGB1 may not be mainly occurring at 6-24 hrs after BNCT, although we observed extracellular release of HMGB1 in culture supernatant with ELISA as previously reported [1].
The limitation of our study is that we have used only two cell lines of oral cancer SAS cells and melanoma A375 cells and we could limitedly analyze an early response after BNCT. For generalization of our results, it may be necessary to analyze using different types of cancer cells with longer observation period. It may be also necessary to analyze using in vivo animal models or clinical samples of patients to understand the biological significances of HMGB1 in BNCT as a therapeutic biomarker.
HMGB1 is reported to be released during necrotic cell death after UV irradiation and gamma irradiation as well as treatment with cytotoxic drugs [5,13,19]. HMGB1 is an abundant and ubiquitously expressed non-histone chromatin protein. It has DNA binding domains and potential to bind minor groove of DNA [3] and acts as a transcription factor and DNA repair factor and is exclusively located in cellular nuclei.
During necrotic cell death, various types of proteins and molecules are released from the cells, which are known as a Damage Associated Molecular Patterns (DAMPs). HMGB1 is known as one of the major DAMPs molecules but the current knowledge is still limited on the functions and roles of DAMPs. The extracellular release of DAMPs is suggested to provoke the alerts to tissue damage in local and systemic manners.
After UV irradiation, the released acetylated HMGB1 binds to the Receptor for Advanced Glycation End products (RAGE) and induces NF-κB and IRF3 mediated transcription of PD-L1 in melanocytes and suppresses the CD8+ T cell-dependent cytotoxicity to melanoma cells [20]. In this process, p38, casein kinase2 and TAK1 are not involved, while RAGE activates TBK1 and phosphorylation of IKKβ further causes phosphorylation of IκB and its degradation and NF-κB activation. On the other hand, He et al. showed after X-ray irradiation, released HMGB1 bind to RAGE, that activates ERK and p38 and promotes cell proliferation [20]. In the case of acetaminophen hepatotoxicity, HMGB1 release is reported to stimulate necrosis by activating RIPK3-dependent pathway and acts on paracrine manner thorough TLK4-TRIF cascade [21]. We have not analyzed which signal transduction pathways are induced during the early responses after BNCT and it is also an open question how local and systemic immune and inflammatory responses are affected by the early HMGB1 extracellular release after neutron beam irradiation with BPA.
On the other hand, HMGB1 is also known to be actively secreted from macrophages, dendritic cells and natural killer cells and other immune cells as an inflammatory response [22]. It is yet to be studied whether macrophages, dendritic cells and natural killer cells actively release HMGB1 after neutron beam irradiation with BPA/γ-irradiation or not.
Cells in dying process are reported to promote repopulation of the tumors after photon beam irradiation [23]. Various cell death processes were shown to be involved in tumor repopulation [19]. It was observed that SAS cells mainly show apoptosis with caspase 3-activation and PARP-1 cleavage during BNCT. We also observed necrotic PARP1 cleavage after 24 therapeutic dose irradiation of BNCT [16]. HMGB1 extracellular release could be possibly induced by the secondary necrosis by shifting from apoptosis in the severely damaged cells. It is also reported that BNCT may directly induce both apoptosis and necrosis [24].
Conclusion
HMGB1 state is mainly regulated at mRNA level but may not be regulated at protein modification levels at early period after BNCT at least in SAS and A375 cells.
Declarations
Funding sources: This work was supported in part by JSPS Grant-in-Aid for Scientific Research, grant numbers 17K15814 (SI) and 18H02693 (MM)) and National Cancer Center Research and Development Fund, grant numbers H26-A-18, H29-A-8 and by research funds from Cancer Intelligence Care Systems, Inc (MM).
Acknowledgements: We thank the kind supports and help of Sota Kikuhara, Hiroaki Fujimori, Soichiro Saito, Takahisa Hirai and the staff members of Kyoto University Research Reactor.
Conflicts of Interest: Research funds from Cancer Intelligence Care Systems, Inc (Mitsuko Masutani).
References
- Imamichi S, Chen L, Ito T, Tong Y, Onodera T, et al. Extracellular release of hmgb1 as an early potential biomarker for the therapeutic response in a xenograft model of boron neutron capture therapy. Biology (Basel). 2022; 11.
- Chiba S, Baghdadi M, Akiba H, Yoshiyama H, Kinoshita I, et al. Tumor-infiltrating dcs suppress nucleic acid-mediated innate immune responses through interactions between the receptor tim-3 and the alarmin hmgb1. Nat Immunol. 2012; 13: 832-842.
- Stott K, Tang GS, Lee KB, Thomas JO. Structure of a complex of tandem hmg boxes and DNA. J Mol Biol. 2006; 360: 90-104.
- Andersson U, Tracey KJ. Hmgb1 is a therapeutic target for sterile inflammation and infection. Annu Rev Immunol. 2011; 29: 139-162.
- Scaffidi P, Misteli T, Bianchi ME. Release of chromatin protein hmgb1 by necrotic cells triggers inflammation. Nature. 2002; 418: 191-195.
- Gardella S, Andrei C, Ferrera D, Lotti LV, Torrisi MR, et al. The nuclear protein hmgb1 is secreted by monocytes via a non-classical, vesicle-mediated secretory pathway. EMBO Rep. 2002; 3: 995-1001.
- Kokkola R, Andersson A, Mullins G, Ostberg T, Treutiger CJ, et al. Rage is the major receptor for the proinflammatory activity of hmgb1 in rodent macrophages. Scand J Immunol. 2005; 61: 1-9.
- Tian J, Avalos AM, Mao SY, Chen B, Senthil K, et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated by hmgb1 and rage. Nat Immunol. 2007; 8: 487-496.
- Zurolo E, Iyer A, Maroso M, Carbonell C, Anink JJ, et al. Activation of toll-like receptor, rage and hmgb1 signalling in malformations of cortical development. Brain. 2011; 134: 1015-1032.
- Masutani M, Baiseitov D, Itoh T, Hirai T, Berikkhanova K, et al. Histological and biochemical analysis of DNA damage after bnct in rat model. Appl Radiat Isot. 2014; 88: 104-108.
- Shi Y, Evans JE, Rock KL. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature. 2003; 425: 516-521.
- Antoine DJ, Jenkins RE, Dear JW, Williams DP, McGill MR, et al. Molecular forms of hmgb1 and keratin-18 as mechanistic biomarkers for mode of cell death and prognosis during clinical acetaminophen hepatotoxicity. J Hepatol. 2012; 56: 1070-1079.
- Balkwill F. Tumour necrosis factor and cancer. Nat Rev Cancer. 2009; 9: 361-371.
- Green DR. The end and after: How dying cells impact the living organism. Immunity. 2011; 35: 441-444.
- Kinashi Y, Takahashi S, Kashino G, Okayasu R, Masunaga S, et al. DNA double-strand break induction in ku80-deficient cho cells following boron neutron capture reaction. Radiat Oncol. 2011; 6: 106.
- Sato A, Itoh T, Imamichi S, Kikuhara S, Fujimori H, et al. Proteomic analysis of cellular response induced by boron neutron capture reaction in human squamous cell carcinoma sas cells. Appl Radiat Isot. 2015; 106: 213-219.
- Yoshino K, Suzuki A, Mori Y, Kakihana H, Honda C, et al. Improvement of solubility of p-boronophenylalanine by complex formation with monosaccharides. Strahlentherapie und Onkologie: Organ der Deutschen Rontgengesellschaft, et al. 1989; 165: 127-129.
- Ditsworth D, Zong WX, Thompson CB. Activation of poly(adp)-ribose polymerase (parp-1) induces release of the pro-inflammatory mediator hmgb1 from the nucleus. J Biol Chem. 2007; 282: 17845-17854.
- He S, Cheng J, Sun L, Wang Y, Wang C, et al. Hmgb1 released by irradiated tumor cells promotes living tumor cell proliferation via paracrine effect. Cell Death Dis. 2018; 9: 648.
- Wang W, Chapman NM, Zhang B, Li M, Fan M, et al. Upregulation of pd-l1 via hmgb1-activated irf3 and nf-kappab contributes to uv radiation- induced immune suppression. Cancer Res. 2019; 79: 2909-2922.
- Minsart C, Liefferinckx C, Lemmers A, Dressen C, Quertinmont E, et al. New insights in acetaminophen toxicity: Hmgb1 contributes by itself to amplify hepatocyte necrosis in vitro through the tlr4-trif-ripk3 axis. Sci Rep. 2020; 10: 5557.
- Williams CD, Bajt ML, Sharpe MR, McGill MR, Farhood A, et al. Neutrophil activation during acetaminophen hepatotoxicity and repair in mice and humans. Toxicol Appl Pharmacol. 2014; 275: 122-133.
- Withers HR, Taylor JM, Maciejewski B. The hazard of accelerated tumor clonogen repopulation during radiotherapy. Acta Oncol. 1988; 27: 131-146.
- Takata T, Kondo N, Sakurai Y, Tanaka H, Hasegawa T, et al. Localized dose delivering by ion beam irradiation for experimental trial of establishing brain necrosis model. Appl Radiat Isot. 2015; 105: 32-34.